

Cysteine and Aspartyl Proteases Contribute to Protein Digestion in the Gut of Freshwater Planaria
- PMID: 27501047
- PMCID: PMC4976874
- DOI: 10.1371/journal.pntd.0004893
Cysteine and Aspartyl Proteases Contribute to Protein Digestion in the Gut of Freshwater Planaria
Abstract
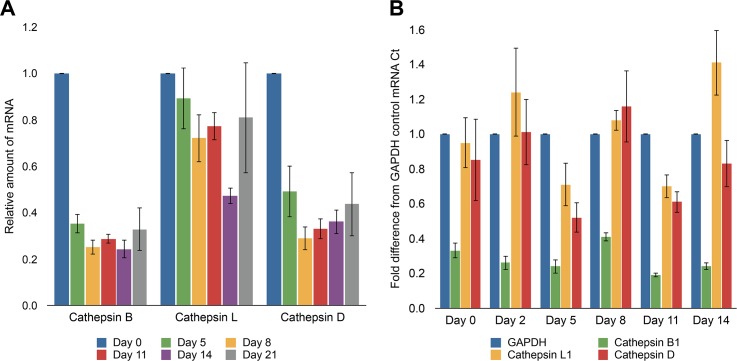
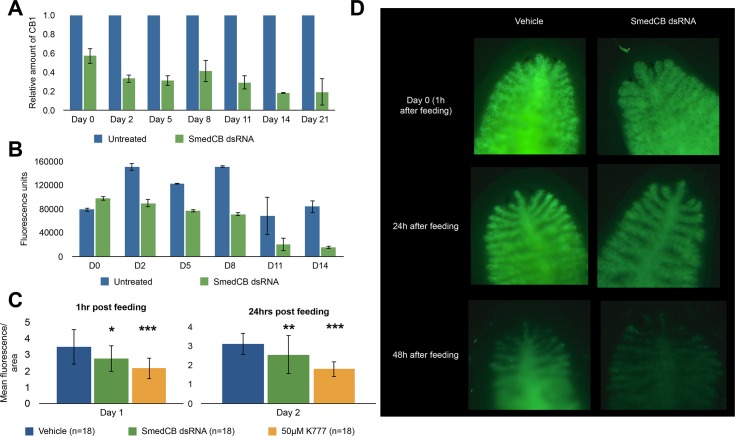
Proteases perform numerous vital functions in flatworms, many of which are likely to be conserved throughout the phylum Platyhelminthes. Within this phylum are several parasitic worms that are often poorly characterized due to their complex life-cycles and lack of responsiveness to genetic manipulation. The flatworm Schmidtea mediterranea, or planaria, is an ideal model organism to study the complex role of protein digestion due to its simple life cycle and amenability to techniques like RNA interference (RNAi). In this study, we were interested in deconvoluting the digestive protease system that exists in the planarian gut. To do this, we developed an alcohol-induced regurgitation technique to enrich for the gut enzymes in S. mediterranea. Using a panel of fluorescent substrates, we show that this treatment produces a sharp increase in proteolytic activity. These enzymes have broad yet diverse substrate specificity profiles. Proteomic analysis of the gut contents revealed the presence of cysteine and metallo-proteases. However, treatment with class-specific inhibitors showed that aspartyl and cysteine proteases are responsible for the majority of protein digestion. Specific RNAi knockdown of the cathepsin B-like cysteine protease (SmedCB) reduced protein degradation in vivo. Immunohistochemistry and whole-mount in situ hybridization (WISH) confirmed that the full-length and active forms of SmedCB are found in secretory cells surrounding the planaria intestinal lumen. Finally, we show that the knockdown of SmedCB reduces the speed of tissue regeneration. Defining the roles of proteases in planaria can provide insight to functions of conserved proteases in parasitic flatworms, potentially uncovering drug targets in parasites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
A Novel Taxon of Monosegmented Double-Stranded RNA Viruses Endemic to Triclad Flatworms.J Virol. 2020 Oct 27;94(22):e00623-20. doi: 10.1128/JVI.00623-20. Print 2020 Oct 27. J Virol. 2020. PMID: 32907972 Free PMC article.
-
Toxoplasma Cathepsin Protease B and Aspartyl Protease 1 Are Dispensable for Endolysosomal Protein Digestion.mSphere. 2020 Feb 12;5(1):e00869-19. doi: 10.1128/mSphere.00869-19. mSphere. 2020. PMID: 32051238 Free PMC article.
-
A multienzyme network functions in intestinal protein digestion by a platyhelminth parasite.J Biol Chem. 2006 Dec 22;281(51):39316-29. doi: 10.1074/jbc.M607128200. Epub 2006 Oct 6. J Biol Chem. 2006. PMID: 17028179
-
Cysteine proteases as digestive enzymes in parasitic helminths.PLoS Negl Trop Dis. 2018 Aug 23;12(8):e0005840. doi: 10.1371/journal.pntd.0005840. eCollection 2018 Aug. PLoS Negl Trop Dis. 2018. PMID: 30138310 Free PMC article. Review.
-
Histone modifications and regeneration in the planarian Schmidtea mediterranea.Curr Top Dev Biol. 2014;108:71-93. doi: 10.1016/B978-0-12-391498-9.00004-8. Curr Top Dev Biol. 2014. PMID: 24512706 Review.
Cited by
-
Multiplex substrate profiling by mass spectrometry for proteases.Methods Enzymol. 2023;682:375-411. doi: 10.1016/bs.mie.2022.09.009. Epub 2022 Dec 21. Methods Enzymol. 2023. PMID: 36948708 Free PMC article.
-
Investigation of three enzymes and their roles in the embryonic development of parthenogenetic Haemaphysalis longicornis.Parasit Vectors. 2020 Jan 31;13(1):46. doi: 10.1186/s13071-020-3916-7. Parasit Vectors. 2020. PMID: 32005284 Free PMC article.
-
Chemosynthetic symbiont with a drastically reduced genome serves as primary energy storage in the marine flatworm Paracatenula.Proc Natl Acad Sci U S A. 2019 Apr 23;116(17):8505-8514. doi: 10.1073/pnas.1818995116. Epub 2019 Apr 8. Proc Natl Acad Sci U S A. 2019. PMID: 30962361 Free PMC article.
-
Muscle functions as a connective tissue and source of extracellular matrix in planarians.Nat Commun. 2019 Apr 8;10(1):1592. doi: 10.1038/s41467-019-09539-6. Nat Commun. 2019. PMID: 30962434 Free PMC article.
-
A metalloprotease produced by larval Schistosoma mansoni facilitates infection establishment and maintenance in the snail host by interfering with immune cell function.PLoS Pathog. 2018 Oct 29;14(10):e1007393. doi: 10.1371/journal.ppat.1007393. eCollection 2018 Oct. PLoS Pathog. 2018. PMID: 30372490 Free PMC article.
References
-
- Morgan T.H. Experimental studies of the regeneration of Planaria maculata. Arch. Entwm. Org. 1898; 7: 364–397.
-
- Reddien PW, Sánchez Alvarado A. Fundamentals of planarian regeneration. Annu. Rev. Cell. Dev. Biol. 2004; 20: 725–757. - PubMed
-
- Sánchez Alvarado A., Newmark P.A., Robb S. M., Juste R. The Schmidtea mediterranea database as a molecular resource for studying platyhelminthes, stem cells and regeneration. Development. 2002; 129: 5659–5665. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
