

Immunotherapy Expands and Maintains the Function of High-Affinity Tumor-Infiltrating CD8 T Cells In Situ
- PMID: 27503208
- PMCID: PMC5010950
- DOI: 10.4049/jimmunol.1502659
Immunotherapy Expands and Maintains the Function of High-Affinity Tumor-Infiltrating CD8 T Cells In Situ
Abstract
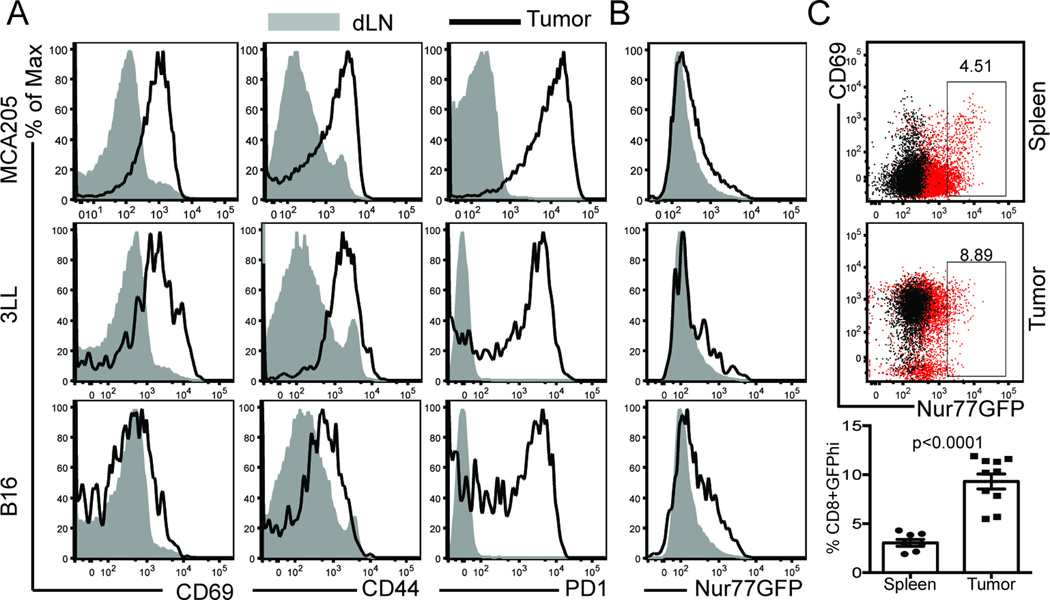
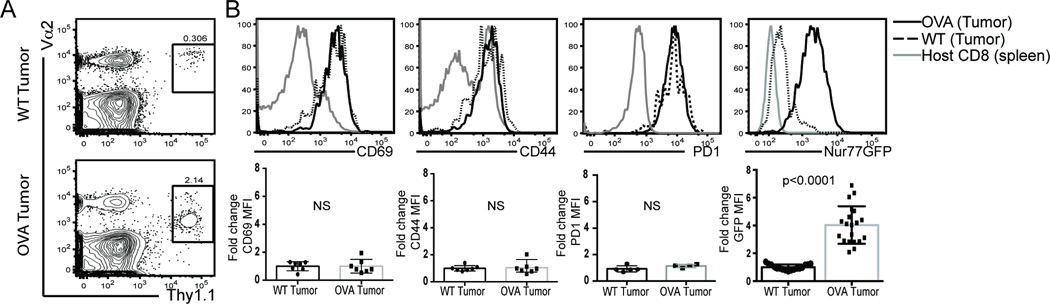
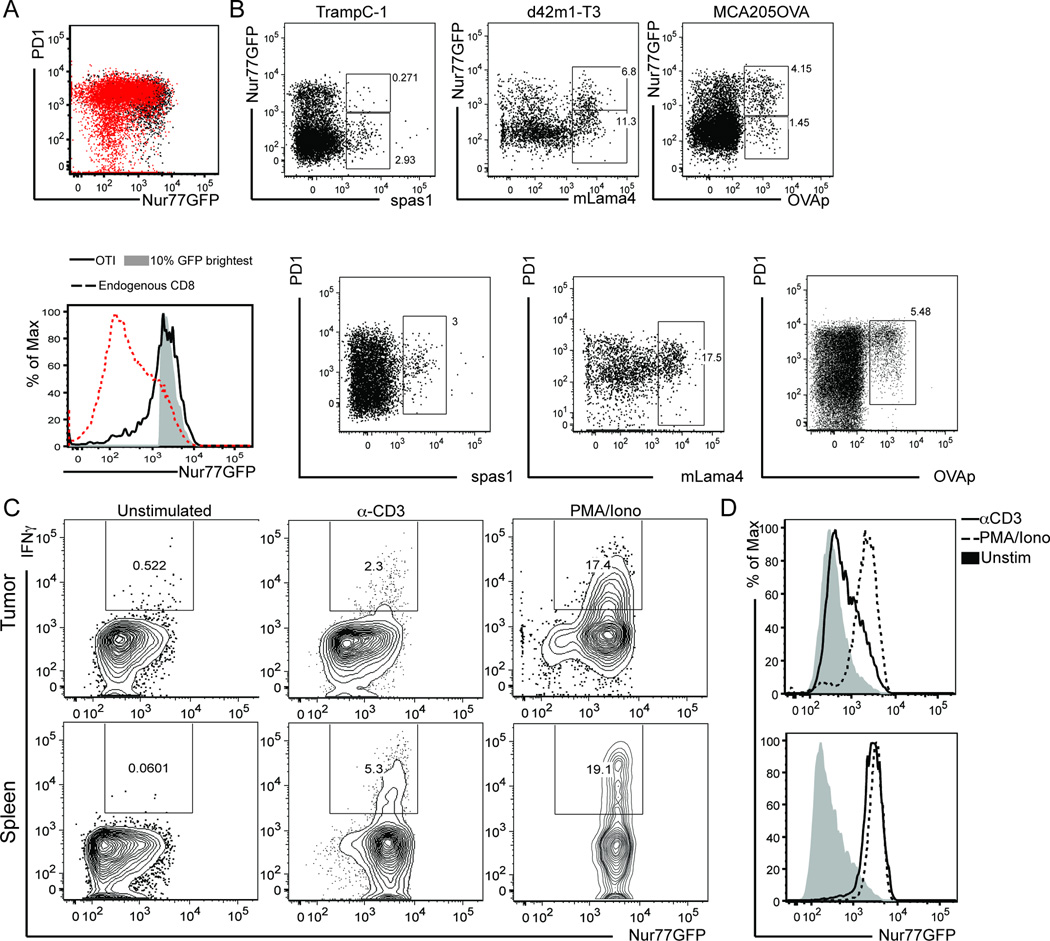
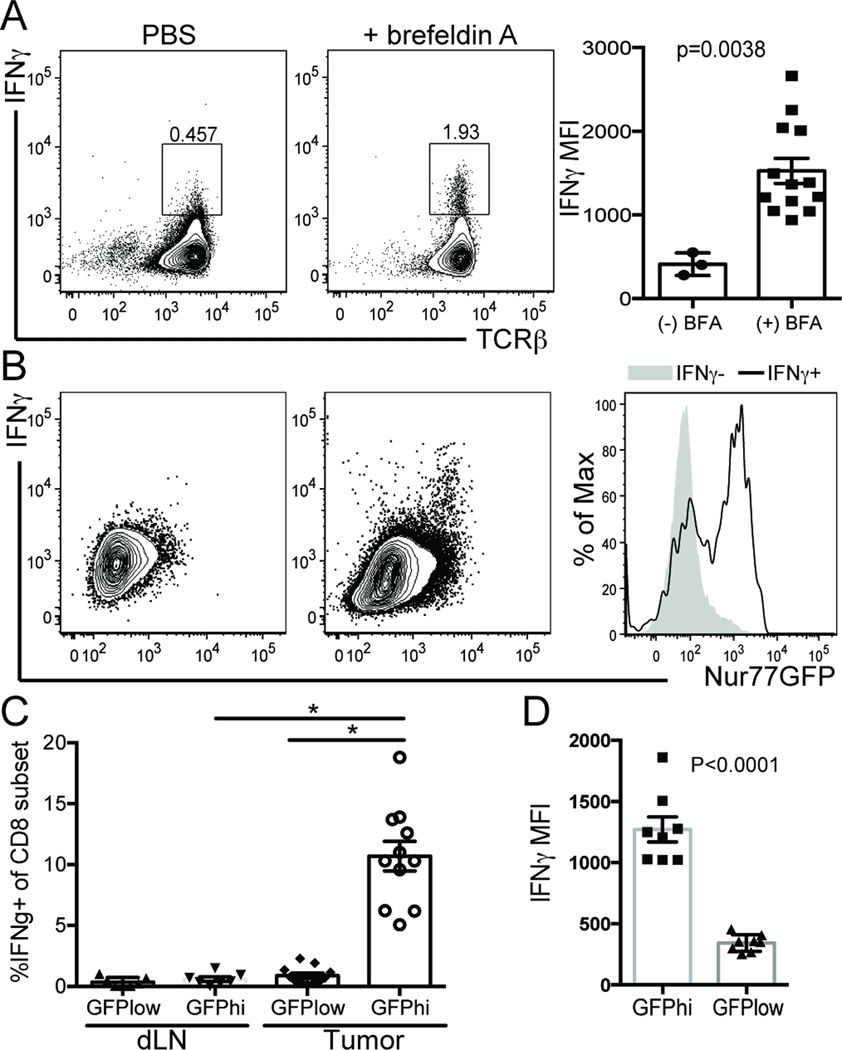
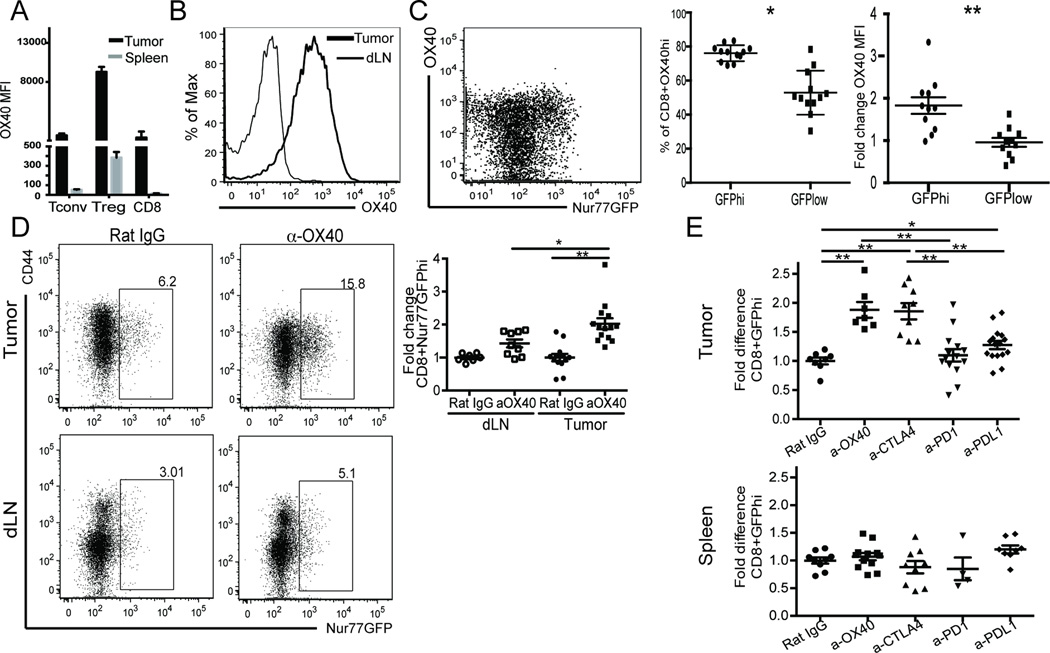
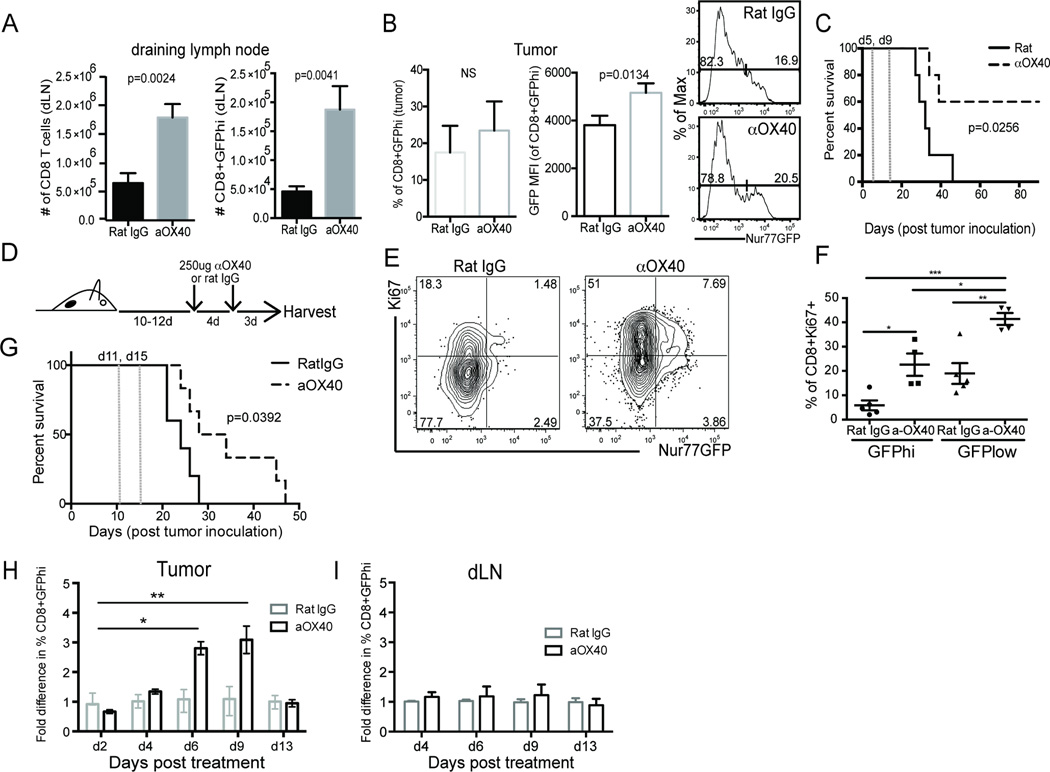
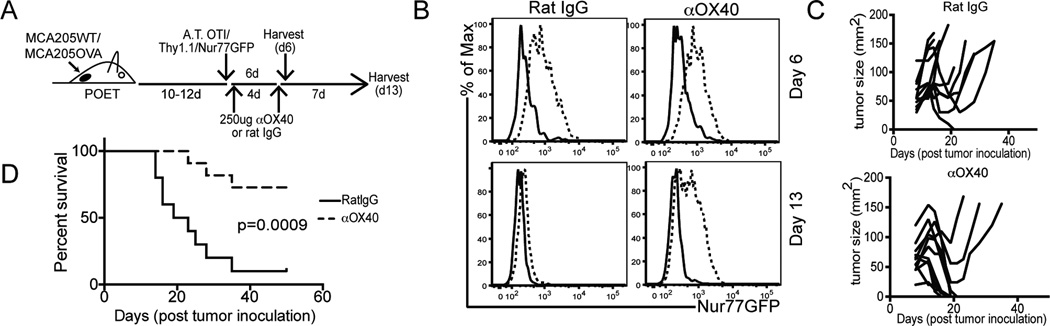
Cancer cells harbor high-affinity tumor-associated Ags capable of eliciting potent antitumor T cell responses, yet detecting these polyclonal T cells is challenging. Therefore, surrogate markers of T cell activation such as CD69, CD44, and programmed death-1 (PD-1) have been used. We report in this study that in mice, expression of activation markers including PD-1 is insufficient in the tumor microenvironment to identify tumor Ag-specific T cells. Using the Nur77GFP T cell affinity reporter mouse, we highlight that PD-1 expression can be induced independent of TCR ligation within the tumor. Given this, we characterized the utility of the Nur77GFP model system in elucidating mechanisms of action of immunotherapies independent of PD-1 expression. Coexpression of Nur77GFP and OX40 identifies a polyclonal population of high-affinity tumor-associated Ag-specific CD8(+) T cells, which produce more IFN-γ in situ than OX40 negative and doubles in quantity with anti-OX40 and anti-CTLA4 mAb therapy but not with anti-PD-1 or programmed death ligand-1. Moreover, expansion of these high-affinity CD8 T cells prolongs survival of tumor-bearing animals. Upon chronic stimulation in tumors and after adoptive cell therapy, CD8 TCR signaling and Nur77GFP induction is impaired, and tumors progress. However, this can be reversed and overall survival significantly enhanced after adoptive cell therapy with agonist OX40 immunotherapy. Therefore, we propose that OX40 agonist immunotherapy can maintain functional TCR signaling of chronically stimulated tumor-resident CD8 T cells, thereby increasing the frequency of cytotoxic, high-affinity, tumor-associated Ag-specific cells.
Copyright © 2016 by The American Association of Immunologists, Inc.
Figures
References
-
- Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Science. 2015;348:69–74. - PubMed
-
- Gubin MM, Zhang X, Schuster H, Caron E, Ward JP, Noguchi T, Ivanova Y, Hundal J, Arthur CD, Krebber WJ, Mulder GE, Toebes M, Vesely MD, Lam SS, Korman AJ, Allison JP, Freeman GJ, Sharpe AH, Pearce EL, Schumacher TN, Aebersold R, Rammensee HG, Melief CJ, Mardis ER, Gillanders WE, Artyomov MN, Schreiber RD. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature. 2014;515:577–581. - PMC - PubMed
-
- Coulie PG, Lehmann F, Lethe B, Herman J, Lurquin C, Andrawiss M, Boon T. A mutated intron sequence codes for an antigenic peptide recognized by cytolytic T lymphocytes on a human melanoma. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:7976–7980. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
