

Differential lipid binding of vinculin isoforms promotes quasi-equivalent dimerization
- PMID: 27503891
- PMCID: PMC5003255
- DOI: 10.1073/pnas.1600702113
Differential lipid binding of vinculin isoforms promotes quasi-equivalent dimerization
Abstract
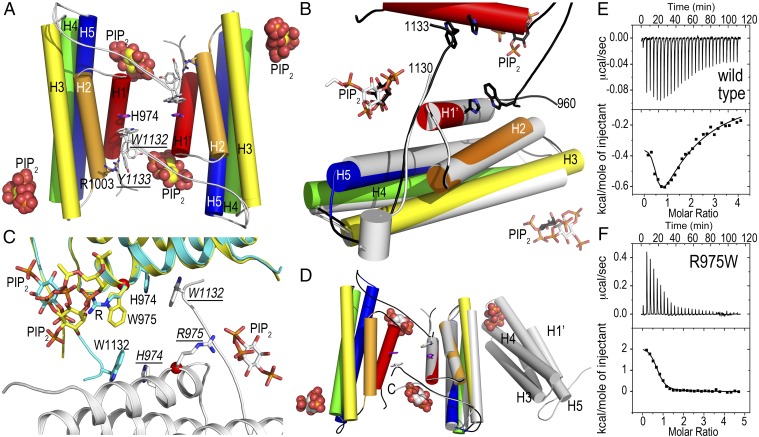
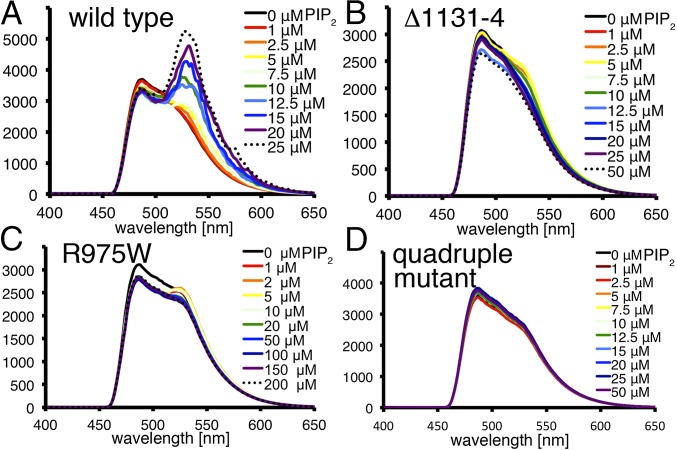
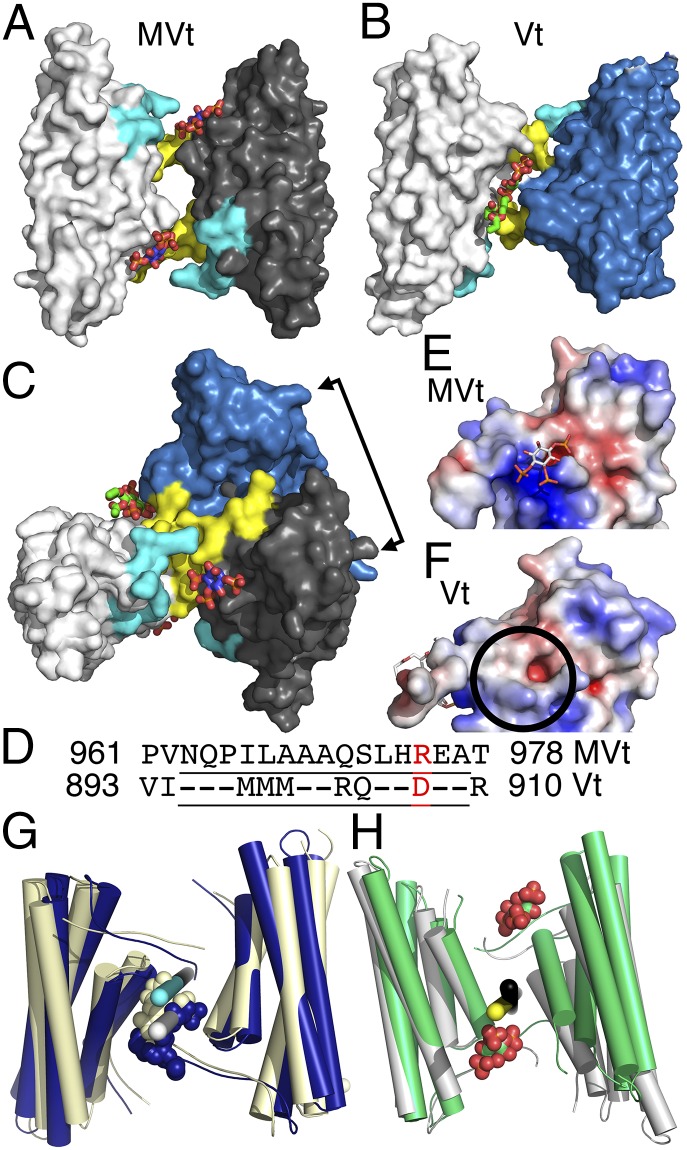
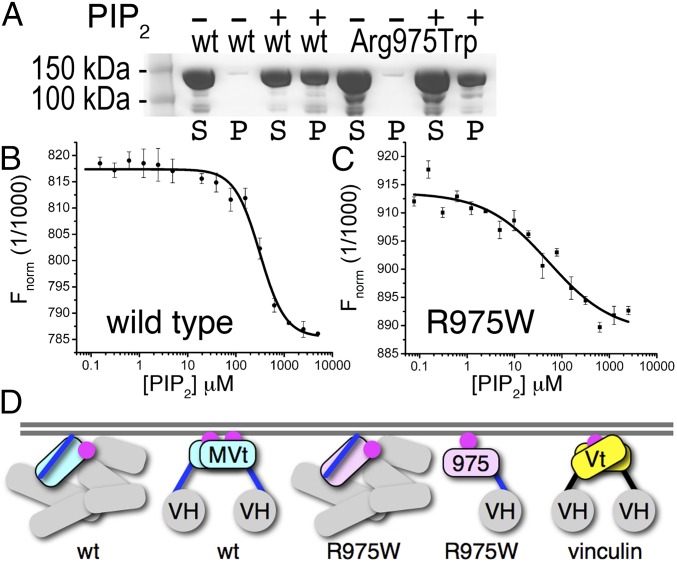
The main cause of death globally remains debilitating heart conditions, such as dilated cardiomyopathy (DCM) and hypertrophic cardiomyopathy (HCM), which are often due to mutations of specific components of adhesion complexes. Vinculin regulates these complexes and plays essential roles in intercalated discs that are necessary for muscle cell function and coordinated movement and in the development and function of the heart. Humans bearing familial or sporadic mutations in vinculin suffer from chronic, progressively debilitating DCM that ultimately leads to cardiac failure and death, whereas autosomal dominant mutations in vinculin can also provoke HCM, causing acute cardiac failure. The DCM/HCM-associated mutants of vinculin occur in the 68-residue insert unique to the muscle-specific, alternatively spliced isoform of vinculin, termed metavinculin (MV). Contrary to studies that suggested that phosphoinositol-4,5-bisphosphate (PIP2) only induces vinculin homodimers, which are asymmetric, we show that phospholipid binding results in a domain-swapped symmetric MV dimer via a quasi-equivalent interface compared with vinculin involving R975. Although one of the two PIP2 binding sites is preserved, the symmetric MV dimer that bridges two PIP2 molecules differs from the asymmetric vinculin dimer that bridges only one PIP2 Unlike vinculin, wild-type MV and the DCM/HCM-associated R975W mutant bind PIP2 in their inactive conformations, and R975W MV fails to dimerize. Mutating selective vinculin residues to their corresponding MV residues, or vice versa, switches the isoform's dimeric constellation and lipid binding site. Collectively, our data suggest that MV homodimerization modulates microfilament attachment at muscular adhesion sites and furthers our understanding of MV-mediated cardiac remodeling.
Keywords: cardiomyopathy; cell adhesion; cytoskeleton; metavinculin; vinculin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Xu W, Baribault H, Adamson ED. Vinculin knockout results in heart and brain defects during embryonic development. Development. 1998;125(2):327–337. - PubMed
-
- Rodríguez Fernández JL, Geiger B, Salomon D, Ben-Ze’ev A. Overexpression of vinculin suppresses cell motility in BALB/c 3T3 cells. Cell Motil Cytoskeleton. 1992;22(2):127–134. - PubMed
-
- Belkin AM, Ornatsky OI, Kabakov AE, Glukhova MA, Koteliansky VE. Diversity of vinculin/meta-vinculin in human tissues and cultivated cells. Expression of muscle specific variants of vinculin in human aorta smooth muscle cells. J Biol Chem. 1988;263(14):6631–6635. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
