

Radial glia in the proliferative ventricular zone of the embryonic and adult turtle, Trachemys scripta elegans
- PMID: 27504470
- PMCID: PMC4973586
- DOI: 10.4161/23262125.2014.970905
Radial glia in the proliferative ventricular zone of the embryonic and adult turtle, Trachemys scripta elegans
Abstract
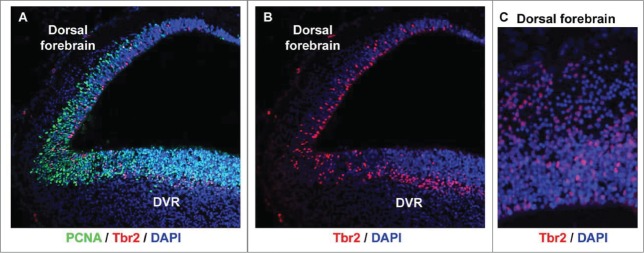
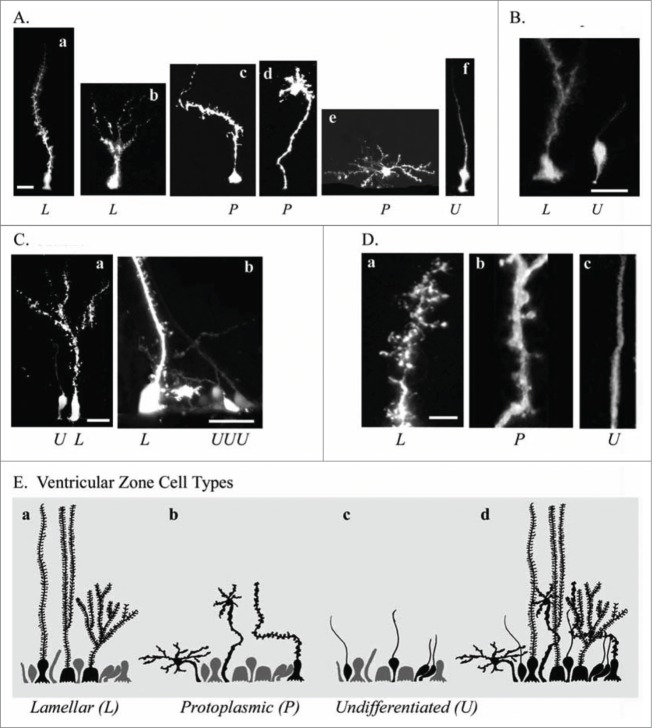
To better understand the role of radial glial (RG) cells in the evolution of the mammalian cerebral cortex, we investigated the role of RG cells in the dorsal cortex and dorsal ventricular ridge of the turtle, Trachemys scripta elegans. Unlike mammals, the glial architecture of adult reptile consists mainly of ependymoradial glia, which share features with mammalian RG cells, and which may contribute to neurogenesis that continues throughout the lifespan of the turtle. To evaluate the morphology and proliferative capacity of ependymoradial glia (here referred to as RG cells) in the dorsal cortex of embryonic and adult turtle, we adapted the cortical electroporation technique, commonly used in rodents, to the turtle telencephalon. Here, we demonstrate the morphological and functional characteristics of RG cells in the developing turtle dorsal cortex. We show that cell division occurs both at the ventricle and away from the ventricle, that RG cells undergo division at the ventricle during neurogenic stages of development, and that mitotic Tbr2+ precursor cells, a hallmark of the mammalian SVZ, are present in the turtle cortex. In the adult turtle, we show that RG cells encompass a morphologically heterogeneous population, particularly in the subpallium where proliferation is most prevalent. One RG subtype is similar to RG cells in the developing mammalian cortex, while 2 other RG subtypes appear to be distinct from those seen in mammal. We propose that the different subtypes of RG cells in the adult turtle perform distinct functions.
Keywords: adult; development; neurogenesis; radial glia; telencephalon; turtle; ventricular zone.
Figures
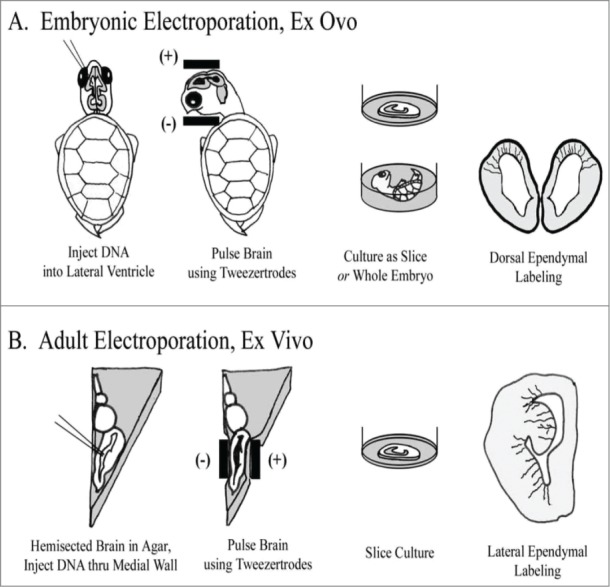
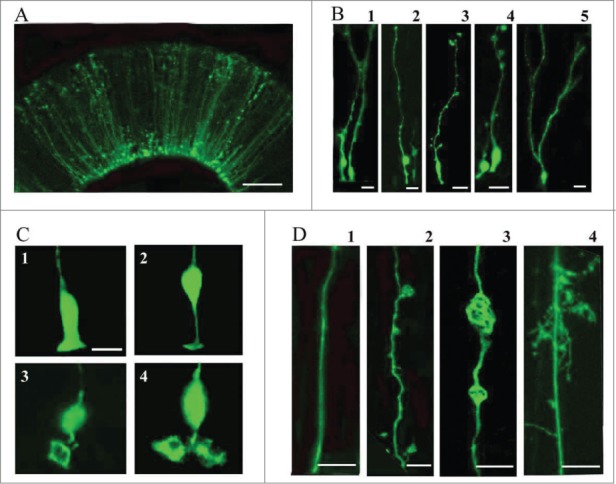
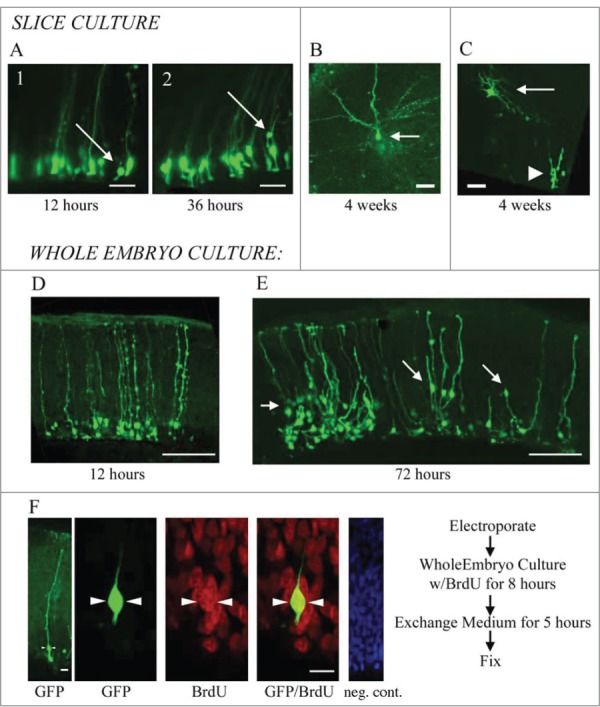
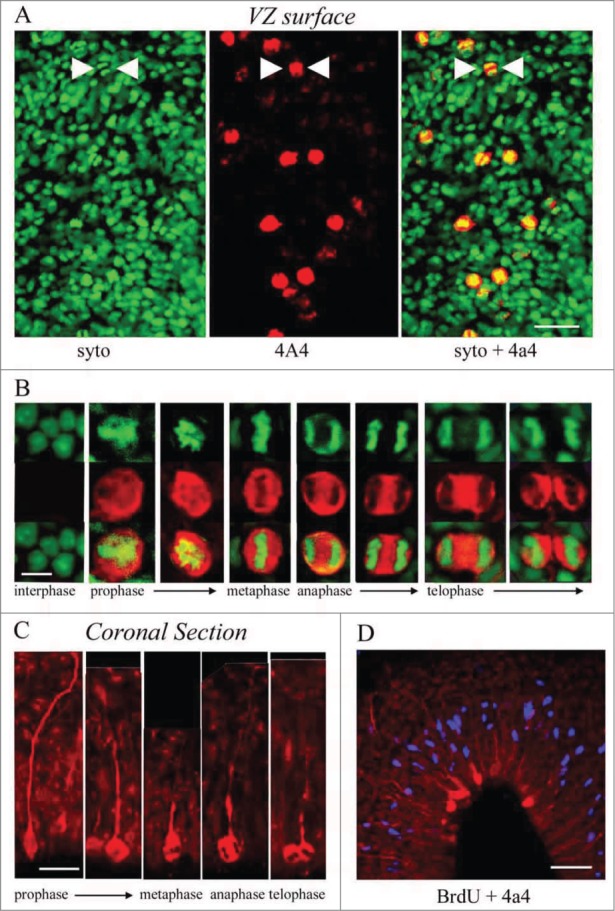
References
-
- Cruce WL, Nieuwenhuys R. The cell masses in the brain stem of the turtle Testudo hermanni; a topographical and topological analysis. J Comp Neurol 1974; 156:277-306; PMID:4418301; http://dx.doi.org/10.1002/cne.901560303 - DOI - PubMed
-
- Blanton MG, Shen JM, Kriegstein AR. Evidence for the inhibitory neurotransmitter gamma-aminobutyric acid in aspiny and sparsely spiny nonpyramidal neurons of the turtle dorsal cortex. J Comp Neurol 1987; 259:277-297; PMID:2438317; http://dx.doi.org/10.1002/cne.902590208 - DOI - PubMed
-
- Levitt P, Rakic P. Immunoperoxidase localization of glial fibrillary acidic protein in radial glial cells and astrocytes of the developing rhesus monkey brain. J Comp Neurol 1980; 193:815-840; PMID:7002963; http://dx.doi.org/10.1002/cne.901930316 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources