

Early programming of the oocyte epigenome temporally controls late prophase I transcription and chromatin remodelling
- PMID: 27507044
- PMCID: PMC4987523
- DOI: 10.1038/ncomms12331
Early programming of the oocyte epigenome temporally controls late prophase I transcription and chromatin remodelling
Abstract
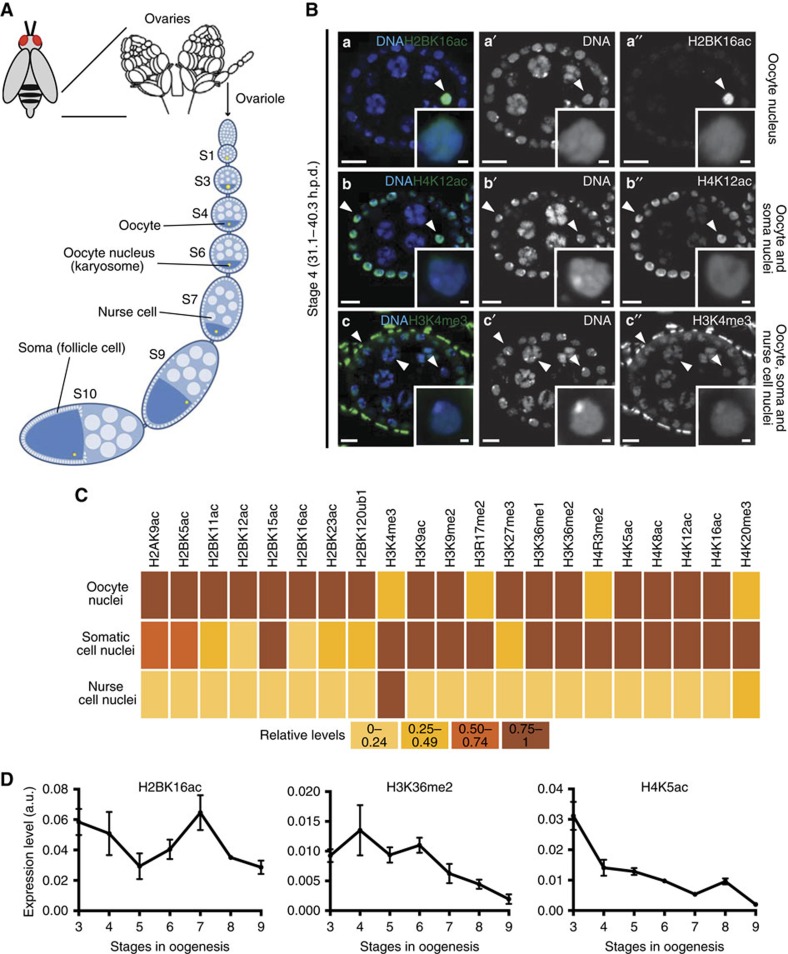
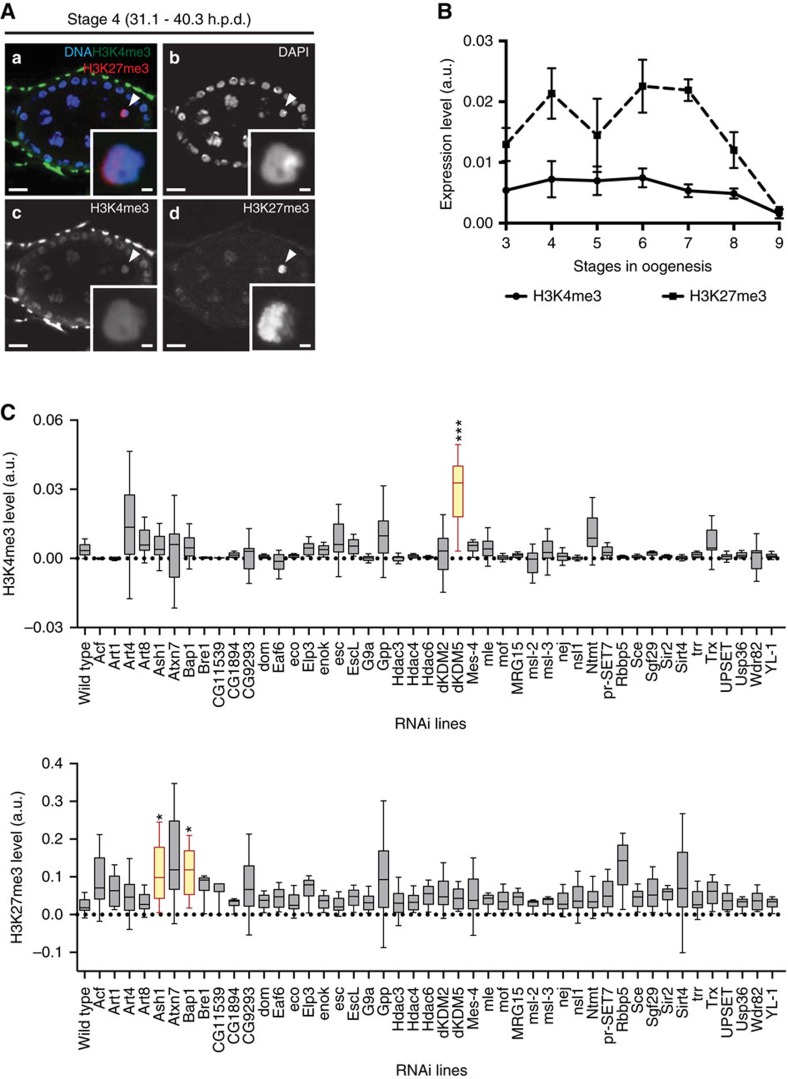
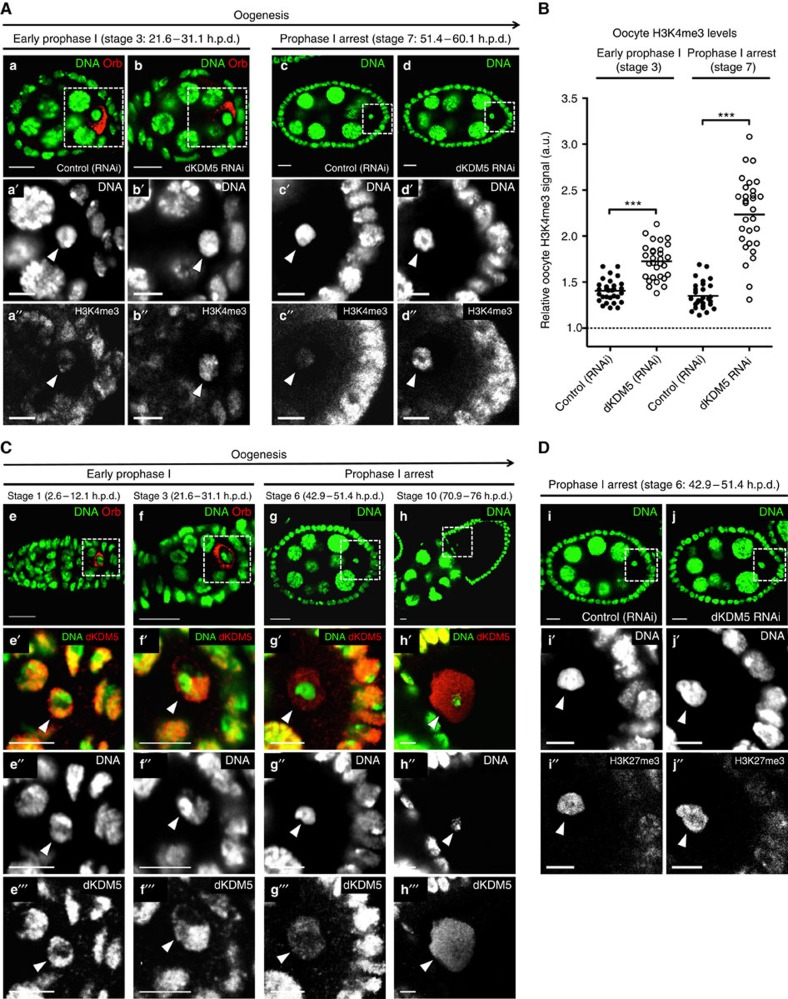
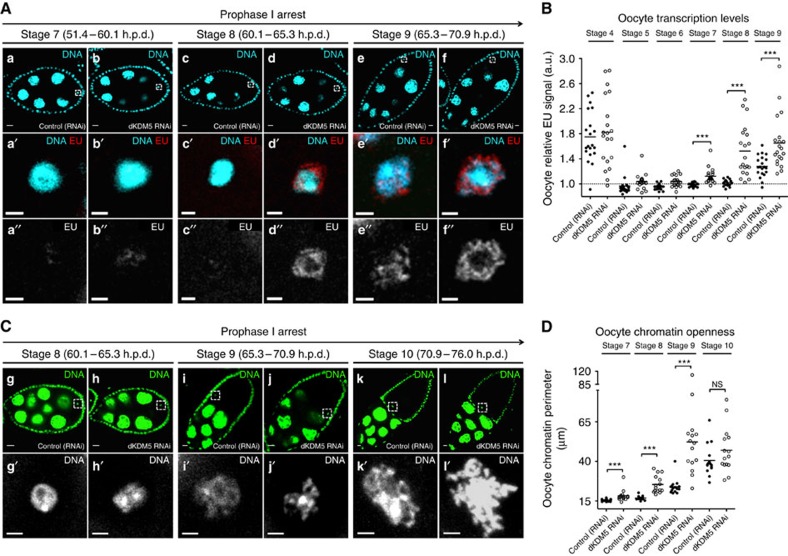
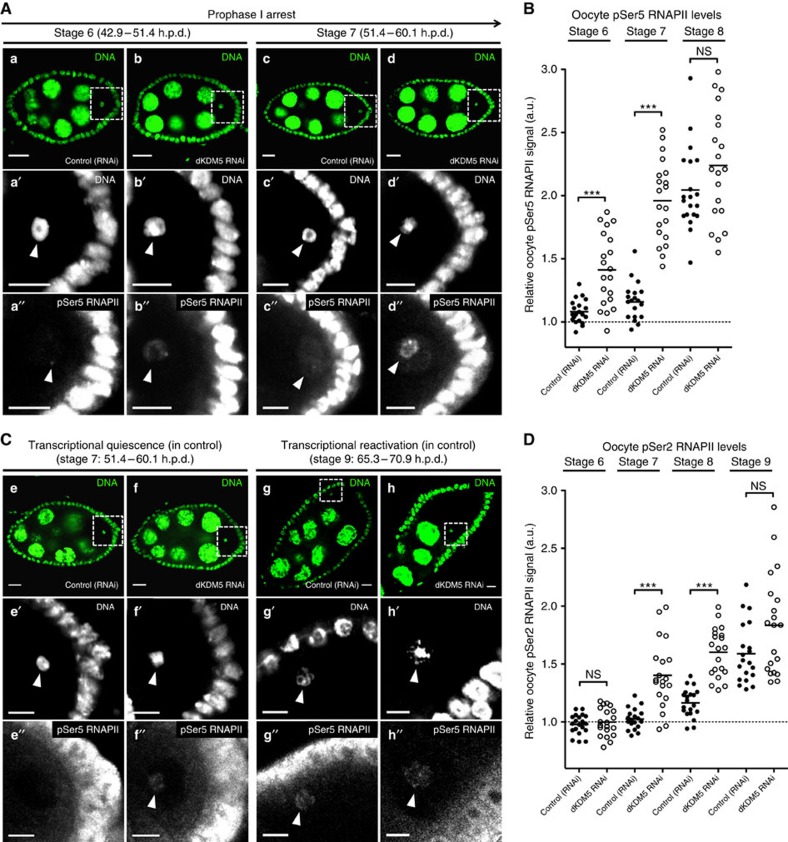
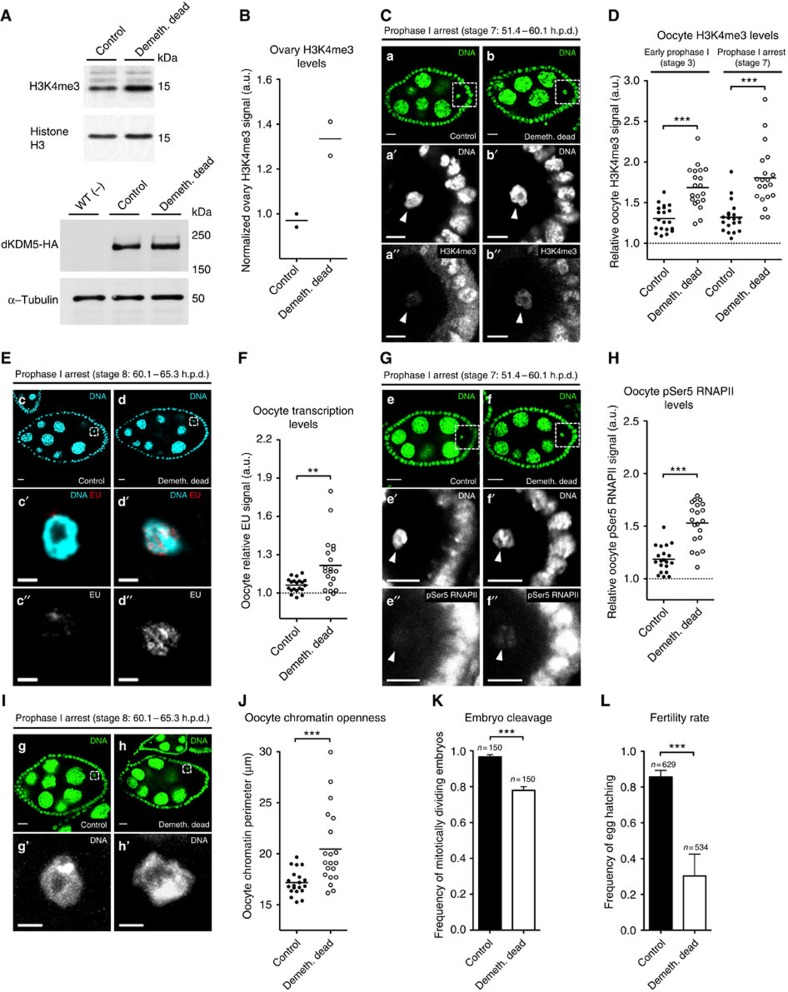
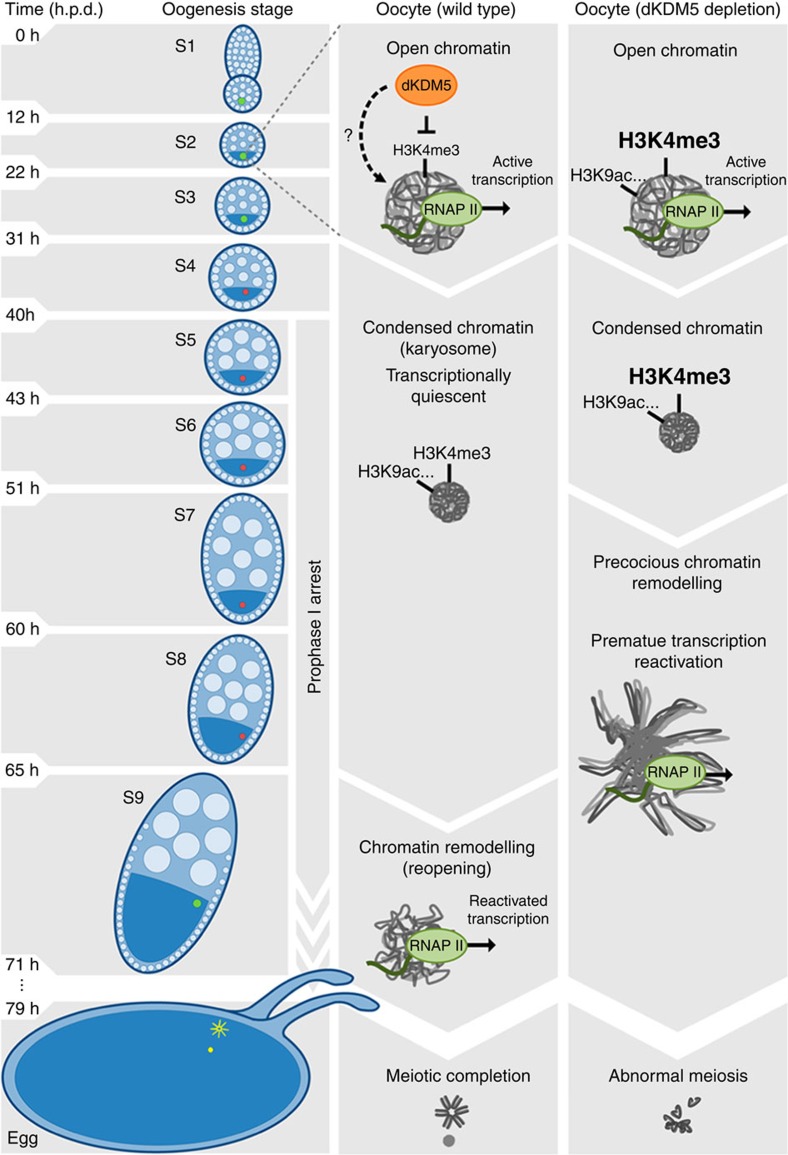
Oocytes are arrested for long periods of time in the prophase of the first meiotic division (prophase I). As chromosome condensation poses significant constraints to gene expression, the mechanisms regulating transcriptional activity in the prophase I-arrested oocyte are still not entirely understood. We hypothesized that gene expression during the prophase I arrest is primarily epigenetically regulated. Here we comprehensively define the Drosophila female germ line epigenome throughout oogenesis and show that the oocyte has a unique, dynamic and remarkably diversified epigenome characterized by the presence of both euchromatic and heterochromatic marks. We observed that the perturbation of the oocyte's epigenome in early oogenesis, through depletion of the dKDM5 histone demethylase, results in the temporal deregulation of meiotic transcription and affects female fertility. Taken together, our results indicate that the early programming of the oocyte epigenome primes meiotic chromatin for subsequent functions in late prophase I.
Figures

References
-
- Gerton J. L. & Hawley R. S. Homologous chromosome interactions in meiosis: diversity amidst conservation. Nat. Rev. Genet. 6, 477–487 (2005). - PubMed
-
- Martinez-Balbas M. A., Dey A., Rabindran S. K., Ozato K. & Wu C. Displacement of sequence-specific transcription factors from mitotic chromatin. Cell 83, 29–38 (1995). - PubMed
-
- Gottesfeld J. M. & Forbes D. J. Mitotic repression of the transcriptional machinery. Trends Biochem. Sci. 22, 197–202 (1997). - PubMed
-
- Davidson E. H. in Gene Activity in Early Development 307–408Academic Press (1986).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
