

RUNX1 contributes to higher-order chromatin organization and gene regulation in breast cancer cells
- PMID: 27514584
- PMCID: PMC5071180
- DOI: 10.1016/j.bbagrm.2016.08.003
RUNX1 contributes to higher-order chromatin organization and gene regulation in breast cancer cells
Abstract
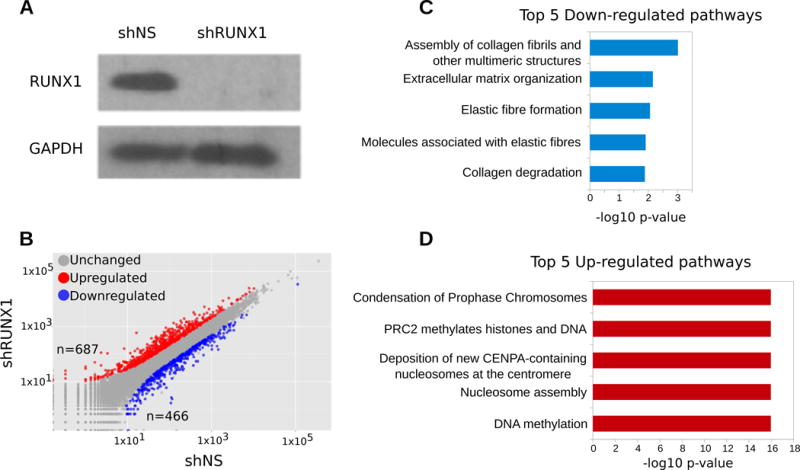
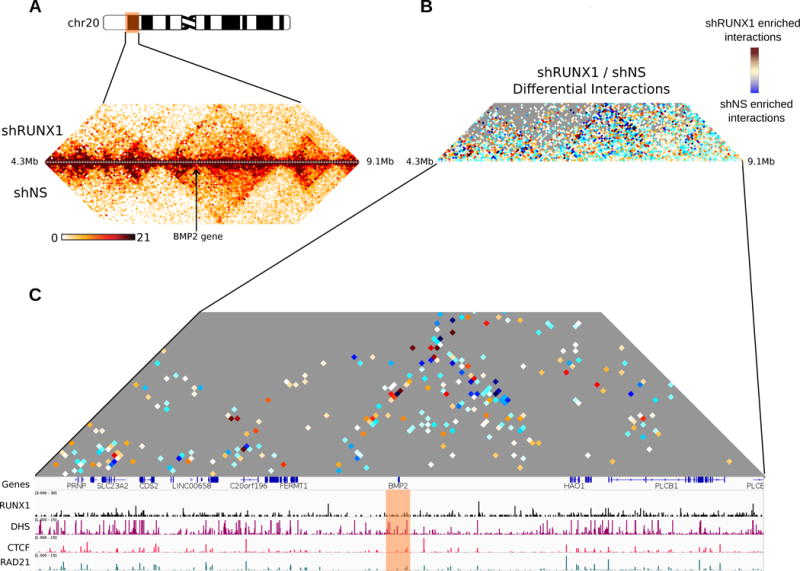
RUNX1 is a transcription factor functioning both as an oncogene and a tumor suppressor in breast cancer. RUNX1 alters chromatin structure in cooperation with chromatin modifier and remodeling enzymes. In this study, we examined the relationship between RUNX1-mediated transcription and genome organization. We characterized genome-wide RUNX1 localization and performed RNA-seq and Hi-C in RUNX1-depleted and control MCF-7 breast cancer cells. RNA-seq analysis showed that RUNX1 depletion led to up-regulation of genes associated with chromatin structure and down-regulation of genes related to extracellular matrix biology, as well as NEAT1 and MALAT1 lncRNAs. Our ChIP-Seq analysis supports a prominent role for RUNX1 in transcriptional activation. About 30% of all RUNX1 binding sites were intergenic, indicating diverse roles in promoter and enhancer regulation and suggesting additional functions for RUNX1. Hi-C analysis of RUNX1-depleted cells demonstrated that overall three-dimensional genome organization is largely intact, but indicated enhanced association of RUNX1 near Topologically Associating Domain (TAD) boundaries and alterations in long-range interactions. These results suggest an architectural role for RUNX1 in fine-tuning local interactions rather than in global organization. Our results provide novel insight into RUNX1-mediated perturbations of higher-order genome organization that are functionally linked with RUNX1-dependent compromised gene expression in breast cancer cells.
Keywords: Breast cancer; Hi-C; MCF-7; RUNX1; TAD; Topologically associating domain.
Copyright © 2016 Elsevier B.V. All rights reserved.
Figures



References
-
- Speck NA, Terryl S. A new transcription factor family associated with human leukemias. Critical reviews in eukaryotic gene expression. 1995;5:337–364. - PubMed
-
- Imperato MR, Cauchy P, Obier N, Bonifer C. The RUNX1-PU.1 axis in the control of hematopoiesis. International journal of hematology. 2015;101:319–329. - PubMed
-
- Lian JB, Stein GS, Javed A, van Wijnen AJ, Stein JL, Montecino M, Hassan MQ, Gaur T, Lengner CJ, Young DW. Networks and hubs for the transcriptional control of osteoblastogenesis. Reviews in endocrine & metabolic disorders. 2006;7:1–16. - PubMed
-
- Chuang LS, Ito K, Ito Y. RUNX family: Regulation and diversification of roles through interacting proteins, International journal of cancer. Journal international du cancer. 2013;132:1260–1271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
