

An intracellular compass spatially coordinates cell cycle modules in Caulobacter crescentus
- PMID: 27517351
- PMCID: PMC5069156
- DOI: 10.1016/j.mib.2016.06.007
An intracellular compass spatially coordinates cell cycle modules in Caulobacter crescentus
Abstract
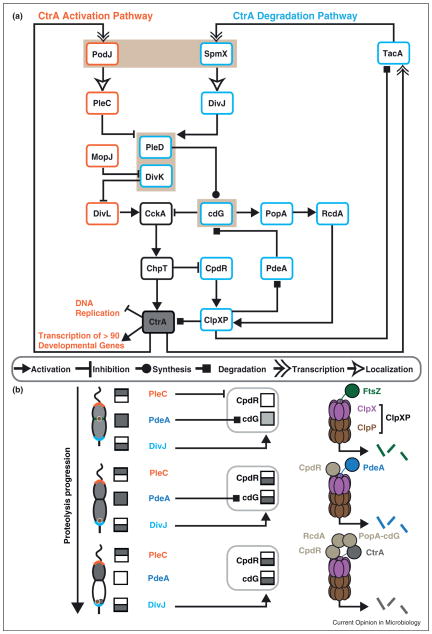
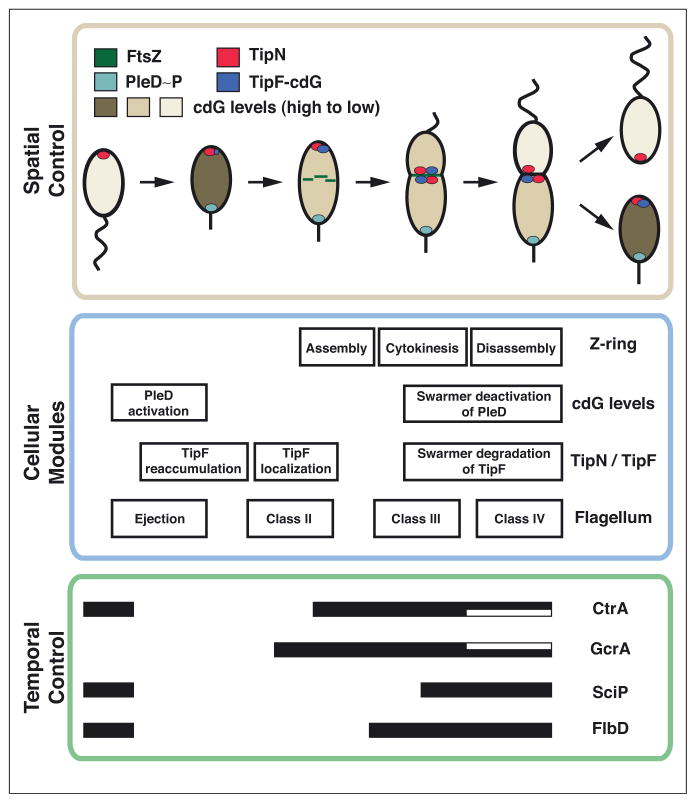
Cellular functions in Bacteria, such as chromosome segregation and cytokinesis, result from cascades of molecular events operating largely as self-contained modules. Regulated timing of these cellular modules stems from global genetic circuits that allow precise temporal activation with respect to cell cycle progression and cell differentiation. Critically, many of these functions occur at defined locations within the cell, and therefore regulators of each module must communicate to remain coordinated in space. In this perspective, we highlight recent discoveries in Caulobacter crescentus asymmetric cell division to illuminate diverse mechanisms by which a cellular compass, composed of scaffolding and signaling proteins, directs cell cycle modules to their exact cellular addresses.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Kirkpatrick CL, Viollier PH. Decoding Caulobacter development. FEMS Microbiol Rev. 2012;36:193–205. - PubMed
-
- Laub MT, McAdams HH, Feldblyum T, Fraser CM, Shapiro L. Global analysis of the genetic network controlling a bacterial cell cycle. Science. 2000;290:2144–2148. - PubMed
-
- Zhou B, Schrader JM, Kalogeraki VS, Abeliuk E, Dinh CB, McAdams HH, Shapiro L. The global regulatory architecture of transcription during the caulobacter cell cycle. PLoS Genet. 2015;11:e1004831. Global analysis of Caulobacter transcription start sites reveals that many cell cycle controlled promoters respond to several of the master regulators. - PMC - PubMed
-
- Panis G, Murray SR, Viollier PH. Versatility of global transcriptional regulators in alpha-Proteobacteria: from essential cell cycle control to ancillary functions. FEMS Microbiol Rev. 2015;39:120–213. - PubMed
-
- Meier EL, Goley ED. Form and function of the bacterial cytokinetic ring. Curr Opin Cell Biol. 2014;26:19–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
