

Chloroquine Analog Interaction with C2- and Iota-Toxin in Vitro and in Living Cells
- PMID: 27517960
- PMCID: PMC4999853
- DOI: 10.3390/toxins8080237
Chloroquine Analog Interaction with C2- and Iota-Toxin in Vitro and in Living Cells
Abstract
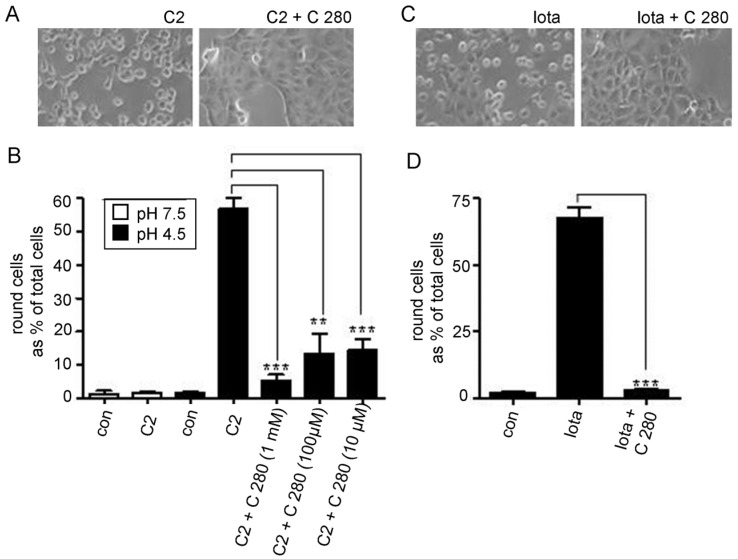
C2-toxin from Clostridium botulinum and Iota-toxin from Clostridium perfringens belong both to the binary A-B-type of toxins consisting of two separately secreted components, an enzymatic subunit A and a binding component B that facilitates the entry of the corresponding enzymatic subunit into the target cells. The enzymatic subunits are in both cases actin ADP-ribosyltransferases that modify R177 of globular actin finally leading to cell death. Following their binding to host cells' receptors and internalization, the two binding components form heptameric channels in endosomal membranes which mediate the translocation of the enzymatic components Iota a and C2I from endosomes into the cytosol of the target cells. The binding components form ion-permeable channels in artificial and biological membranes. Chloroquine and related 4-aminoquinolines were able to block channel formation in vitro and intoxication of living cells. In this study, we extended our previous work to the use of different chloroquine analogs and demonstrate that positively charged aminoquinolinium salts are able to block channels formed in lipid bilayer membranes by the binding components of C2- and Iota-toxin. Similarly, these molecules protect cultured mammalian cells from intoxication with C2- and Iota-toxin. The aminoquinolinium salts did presumably not interfere with actin ADP-ribosylation or receptor binding but blocked the pores formed by C2IIa and Iota b in living cells and in vitro. The blocking efficiency of pores formed by Iota b and C2IIa by the chloroquine analogs showed interesting differences indicating structural variations between the types of protein-conducting nanochannels formed by Iota b and C2IIa.
Keywords: C2-toxin; aminoquinolinium salts; binding components; black lipid bilayer; chloroquine; iota-toxin.
Figures
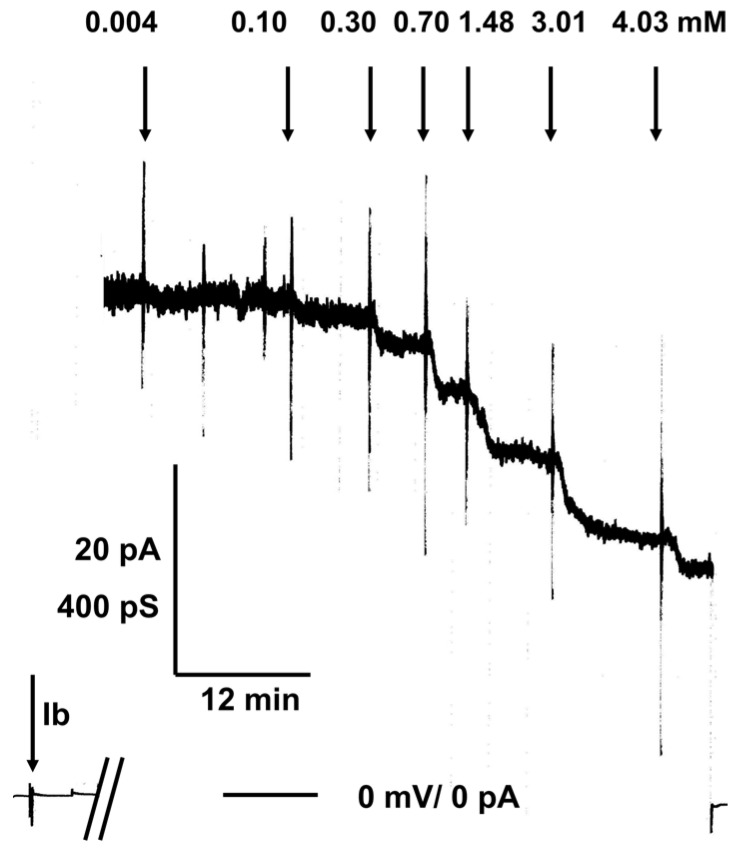
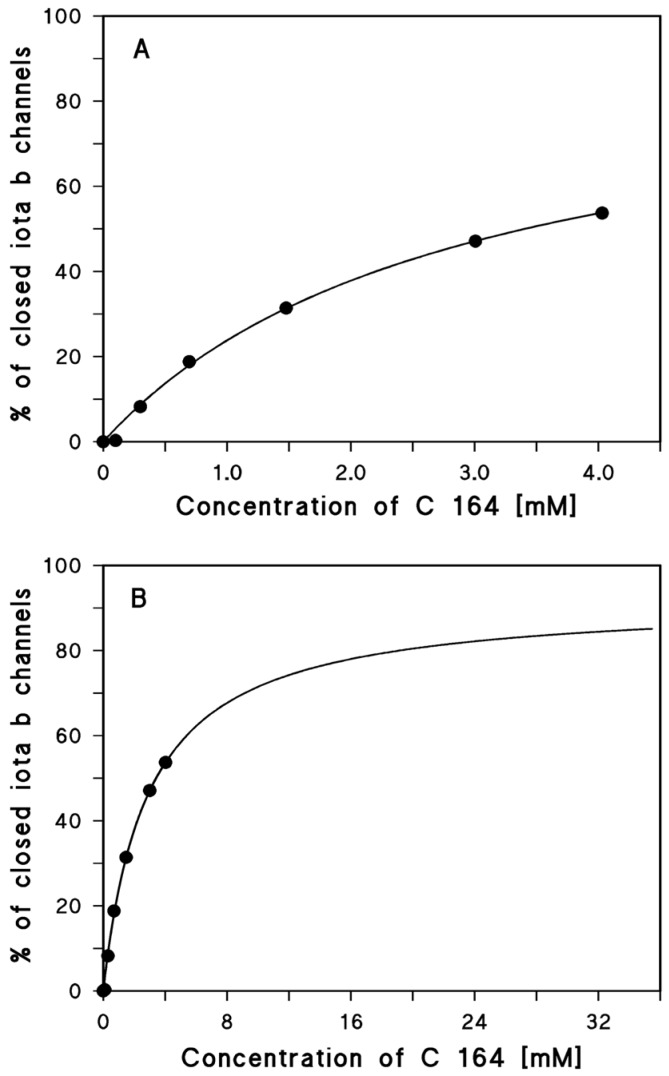
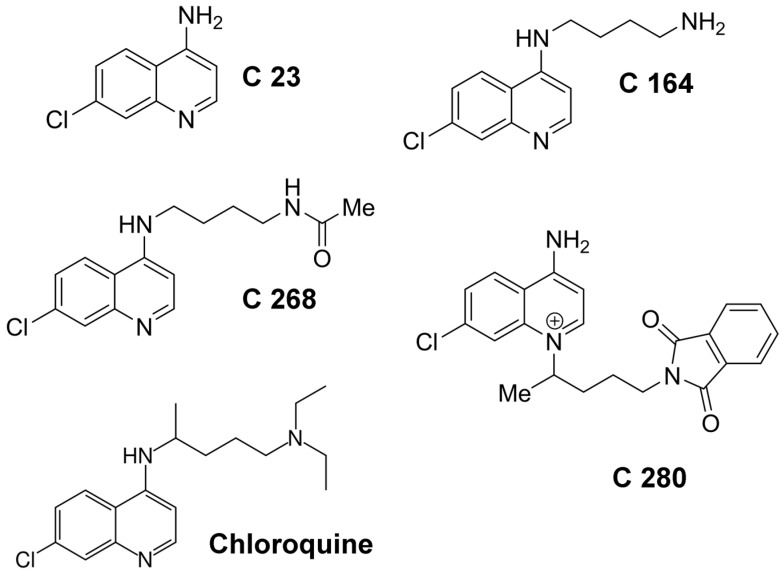
References
-
- Aktories K., Barth H. New insights into the mode of action of the actin ADP-ribosylating virulence factors Salmonella enterica SpvB and Clostridium botulinum C2 toxin. Eur. J. Cell Biol. 2011;90:944–950. - PubMed
-
- Friedlander A.M. Macrophages are sensitive to anthrax lethal toxin through an acid-dependent process. J. Biol. Chem. 1986;261:7123–7126. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
