

Light-Controlled Cell Factories: Employing Photocaged Isopropyl-β-d-Thiogalactopyranoside for Light-Mediated Optimization of lac Promoter-Based Gene Expression and (+)-Valencene Biosynthesis in Corynebacterium glutamicum
- PMID: 27520809
- PMCID: PMC5068161
- DOI: 10.1128/AEM.01457-16
Light-Controlled Cell Factories: Employing Photocaged Isopropyl-β-d-Thiogalactopyranoside for Light-Mediated Optimization of lac Promoter-Based Gene Expression and (+)-Valencene Biosynthesis in Corynebacterium glutamicum
Abstract
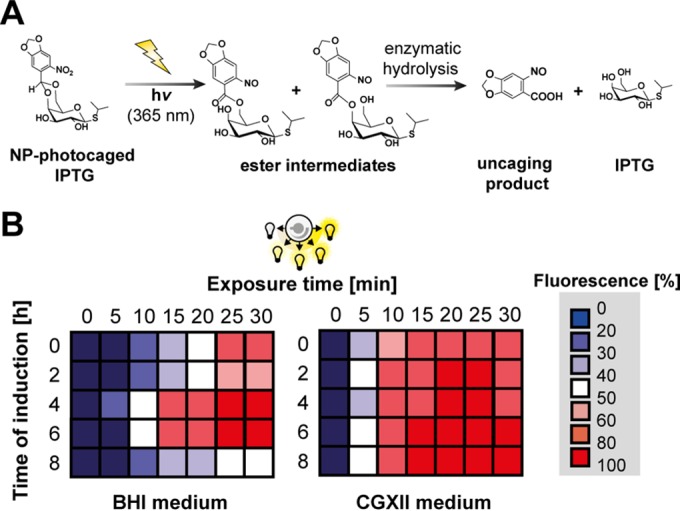
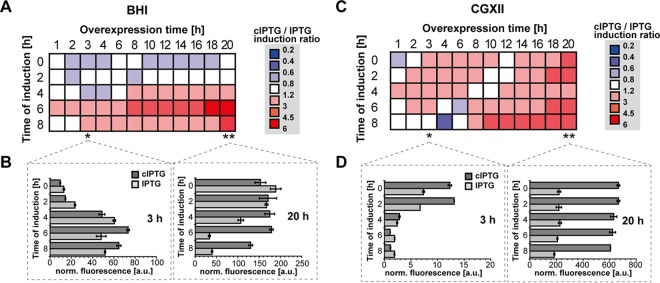
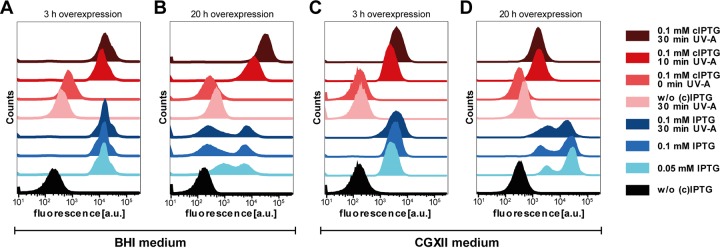
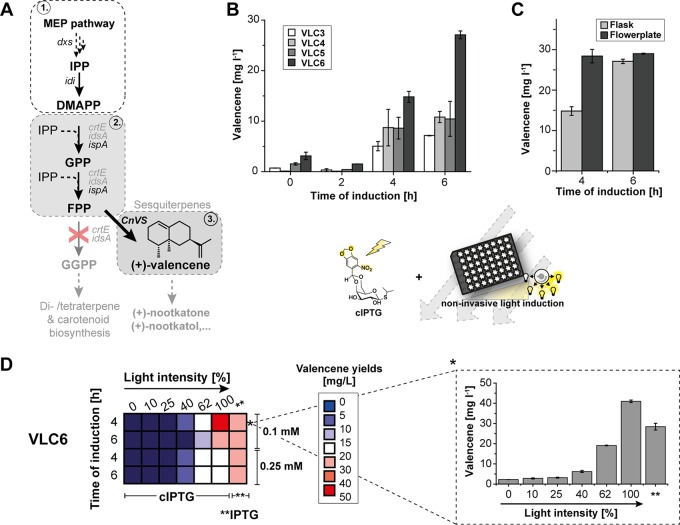
Precise control of microbial gene expression resulting in a defined, fast, and homogeneous response is of utmost importance for synthetic bio(techno)logical applications. However, even broadly applied biotechnological workhorses, such as Corynebacterium glutamicum, for which induction of recombinant gene expression commonly relies on the addition of appropriate inducer molecules, perform moderately in this respect. Light offers an alternative to accurately control gene expression, as it allows for simple triggering in a noninvasive fashion with unprecedented spatiotemporal resolution. Thus, optogenetic switches are promising tools to improve the controllability of existing gene expression systems. In this regard, photocaged inducers, whose activities are initially inhibited by light-removable protection groups, represent one of the most valuable photoswitches for microbial gene expression. Here, we report on the evaluation of photocaged isopropyl-β-d-thiogalactopyranoside (IPTG) as a light-responsive control element for the frequently applied tac-based expression module in C. glutamicum In contrast to conventional IPTG, the photocaged inducer mediates a tightly controlled, strong, and homogeneous expression response upon short exposure to UV-A light. To further demonstrate the unique potential of photocaged IPTG for the optimization of production processes in C. glutamicum, the optogenetic switch was finally used to improve biosynthesis of the growth-inhibiting sesquiterpene (+)-valencene, a flavoring agent and aroma compound precursor in food industry. The variation in light intensity as well as the time point of light induction proved crucial for efficient production of this toxic compound.
Importance: Optogenetic tools are light-responsive modules that allow for a simple triggering of cellular functions with unprecedented spatiotemporal resolution and in a noninvasive fashion. Specifically, light-controlled gene expression exhibits an enormous potential for various synthetic bio(techno)logical purposes. Before our study, poor inducibility, together with phenotypic heterogeneity, was reported for the IPTG-mediated induction of lac-based gene expression in Corynebacterium glutamicum By applying photocaged IPTG as a synthetic inducer, however, these drawbacks could be almost completely abolished. Especially for increasing numbers of parallelized expression cultures, noninvasive and spatiotemporal light induction qualifies for a precise, homogeneous, and thus higher-order control to fully automatize or optimize future biotechnological applications.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Light-responsive control of bacterial gene expression: precise triggering of the lac promoter activity using photocaged IPTG.Integr Biol (Camb). 2014 Aug;6(8):755-65. doi: 10.1039/c4ib00027g. Integr Biol (Camb). 2014. PMID: 24894989
-
Light-induced gene expression with photocaged IPTG for induction profiling in a high-throughput screening system.Microb Cell Fact. 2016 Apr 23;15:63. doi: 10.1186/s12934-016-0461-3. Microb Cell Fact. 2016. PMID: 27107964 Free PMC article.
-
A chromosomally encoded T7 RNA polymerase-dependent gene expression system for Corynebacterium glutamicum: construction and comparative evaluation at the single-cell level.Microb Biotechnol. 2015 Mar;8(2):253-65. doi: 10.1111/1751-7915.12236. Epub 2014 Dec 9. Microb Biotechnol. 2015. PMID: 25488698 Free PMC article.
-
Expression of recombinant protein using Corynebacterium Glutamicum: progress, challenges and applications.Crit Rev Biotechnol. 2016 Aug;36(4):652-64. doi: 10.3109/07388551.2015.1004519. Epub 2015 Feb 25. Crit Rev Biotechnol. 2016. PMID: 25714007 Review.
-
Tools for genetic manipulations in Corynebacterium glutamicum and their applications.Appl Microbiol Biotechnol. 2011 Jun;90(5):1641-54. doi: 10.1007/s00253-011-3272-9. Epub 2011 Apr 26. Appl Microbiol Biotechnol. 2011. PMID: 21519933 Review.
Cited by
-
Engineered Rhodobacter capsulatus as a Phototrophic Platform Organism for the Synthesis of Plant Sesquiterpenoids.Front Microbiol. 2019 Sep 6;10:1998. doi: 10.3389/fmicb.2019.01998. eCollection 2019. Front Microbiol. 2019. PMID: 31555236 Free PMC article.
-
Isoprenoid Pyrophosphate-Dependent Transcriptional Regulation of Carotenogenesis in Corynebacterium glutamicum.Front Microbiol. 2017 Apr 24;8:633. doi: 10.3389/fmicb.2017.00633. eCollection 2017. Front Microbiol. 2017. PMID: 28484430 Free PMC article.
-
Multiplexed characterization of rationally designed promoter architectures deconstructs combinatorial logic for IPTG-inducible systems.Nat Commun. 2021 Jan 12;12(1):325. doi: 10.1038/s41467-020-20094-3. Nat Commun. 2021. PMID: 33436562 Free PMC article.
-
Synthesis and Application of Photocaged Isopropyl β-D-1-Thiogalactopyranoside for Light-Mediated Control of Bacterial Gene Expression.Methods Mol Biol. 2025;2840:133-148. doi: 10.1007/978-1-0716-4047-0_10. Methods Mol Biol. 2025. PMID: 39724349
-
CRISPRi-Library-Guided Target Identification for Engineering Carotenoid Production by Corynebacterium glutamicum.Microorganisms. 2021 Mar 24;9(4):670. doi: 10.3390/microorganisms9040670. Microorganisms. 2021. PMID: 33805131 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
