

Phosphorylation-Induced Motor Shedding Is Required at Mitosis for Proper Distribution and Passive Inheritance of Mitochondria
- PMID: 27524620
- PMCID: PMC5001922
- DOI: 10.1016/j.celrep.2016.07.055
Phosphorylation-Induced Motor Shedding Is Required at Mitosis for Proper Distribution and Passive Inheritance of Mitochondria
Abstract
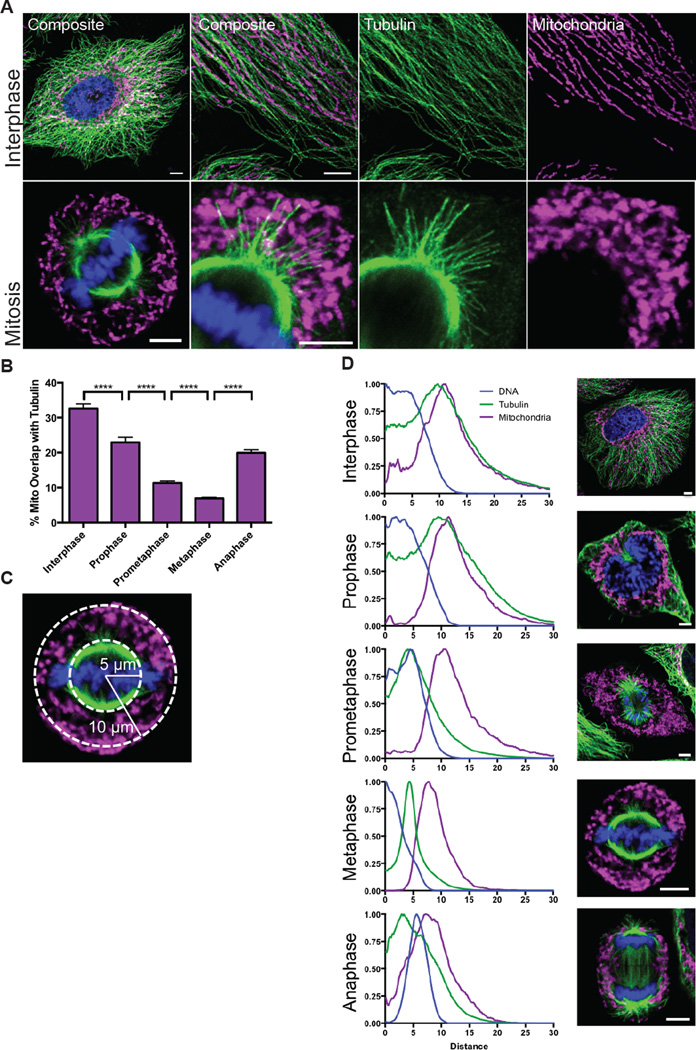
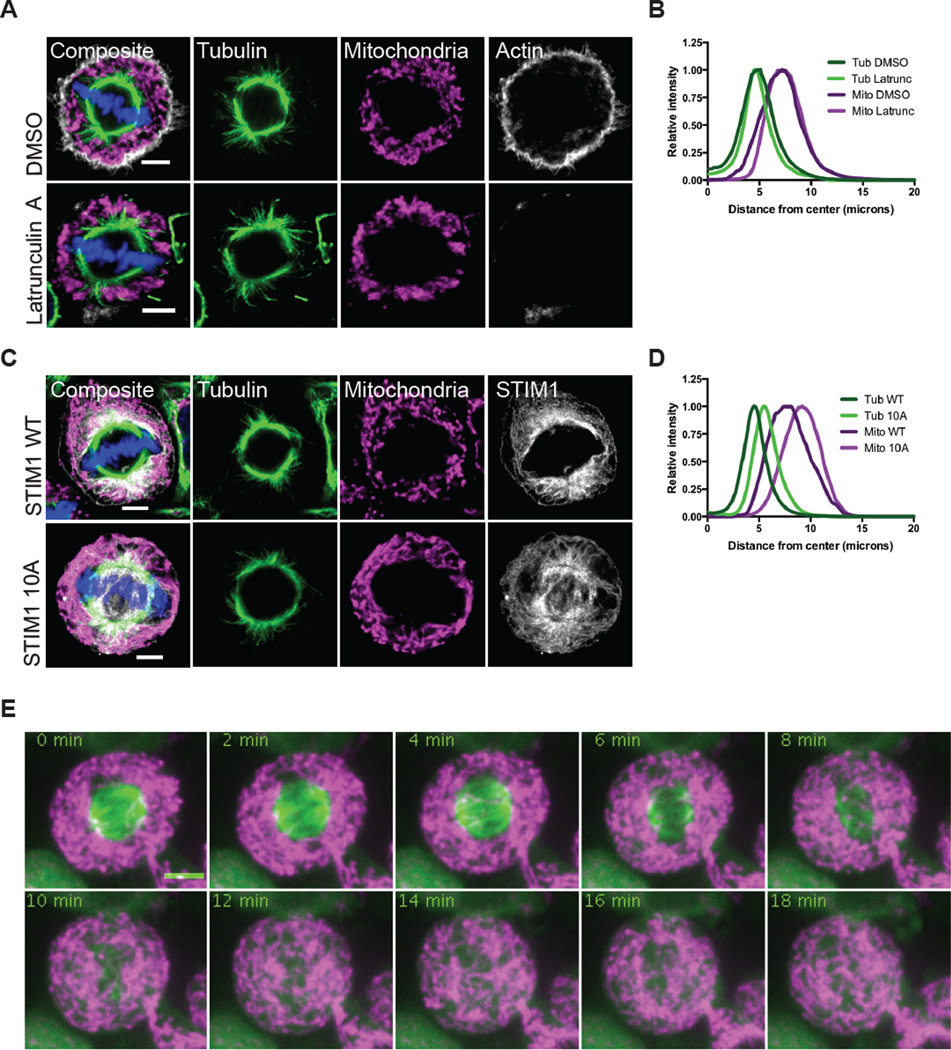
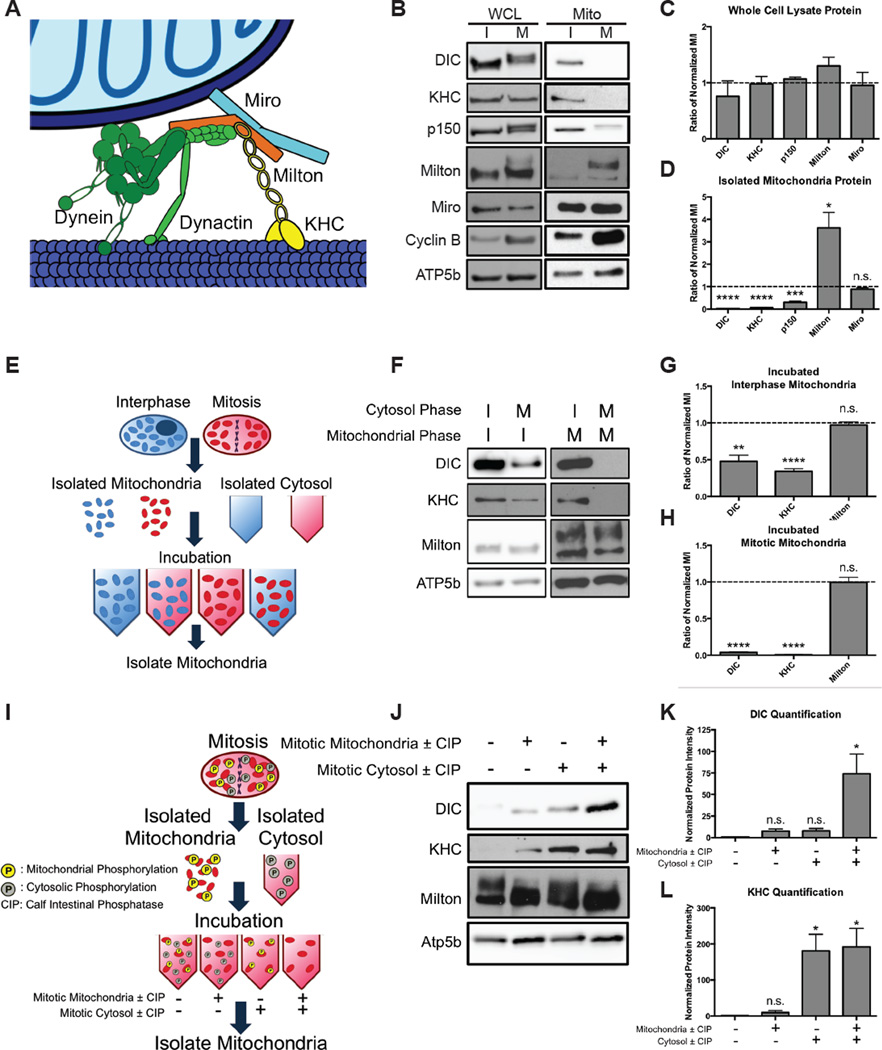
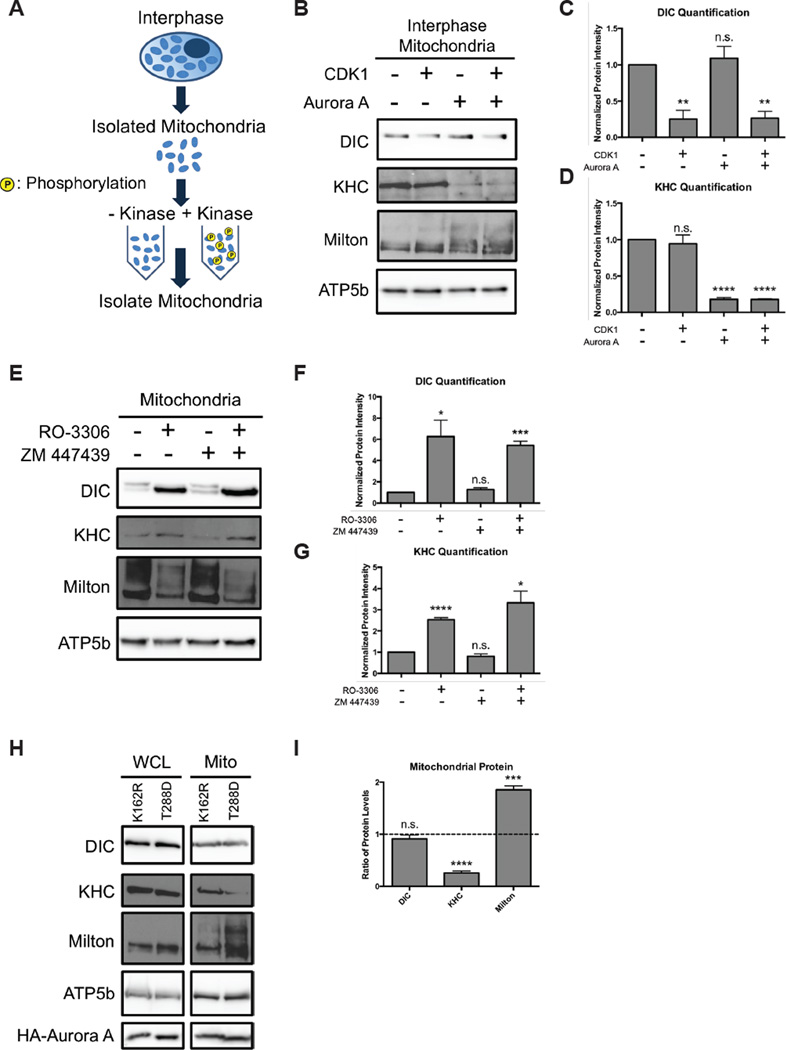
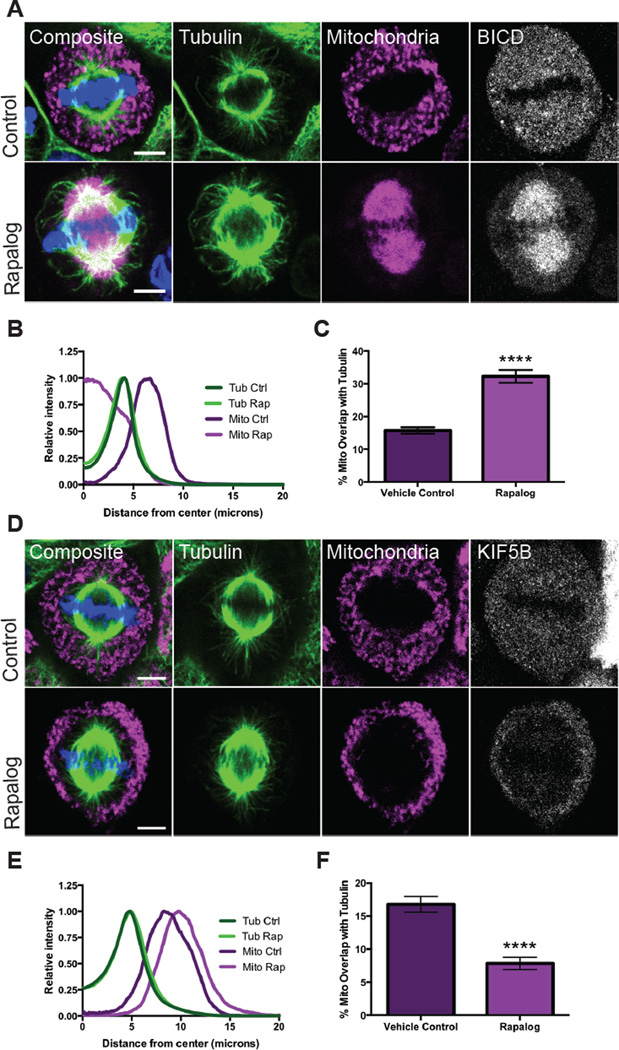
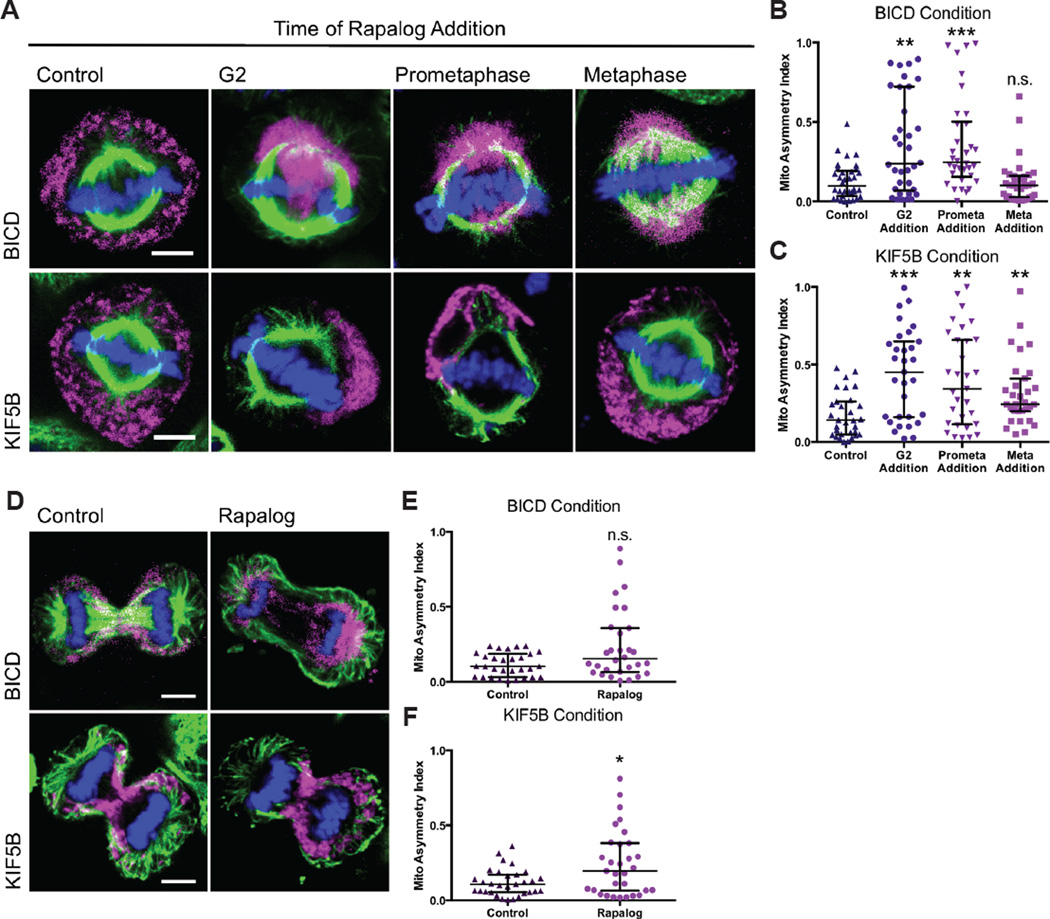
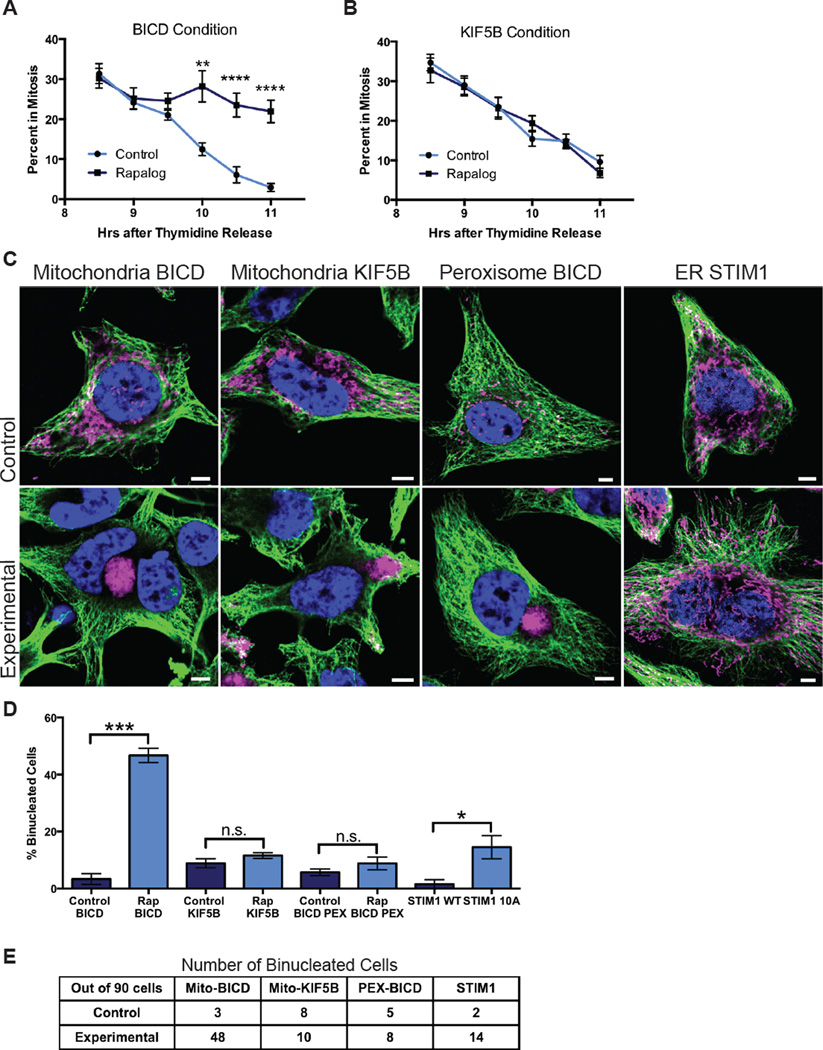
While interphase mitochondria associate with microtubules, mitotic mitochondria dissociate from spindle microtubules and localize in the cell periphery. Here, we show that this redistribution is not mediated by mitochondrial active transport or tethering to the cytoskeleton. Instead, kinesin and dynein, which link mitochondria to microtubules, are shed from the mitochondrial surface. Shedding is driven by phosphorylation of mitochondrial and cytoplasmic targets by CDK1 and Aurora A. Forced recruitment of motor proteins to mitotic mitochondria to override this shedding prevents their proper symmetrical distribution and disrupts the balanced inheritance of mitochondria to daughter cells. Moreover, when mitochondria with bound dynein bind to the mitotic spindle, they arrest cell-cycle progression and produce binucleate cells. Thus, our results show that the regulated release of motor proteins from the mitochondrial surface is a critical mitotic event.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Baggethun P. 2009 Radial Profile Plot ( http://rsb.info.nih.gov/ij/plugins/radial-profile.html: U. S. National I..., Bethesda, Maryland, USA)
-
- de Brito OM, Scorrano L. Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature. 2008;456:605–610. - PubMed
-
- Dell KR, Turck CW, Vale RD. Mitotic phosphorylation of the dynein light intermediate chain is mediated by cdc2 kinase. Traffic (Copenhagen, Denmark) 2000;1:38–44. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
