

p62/Sequestosome-1 Is Indispensable for Maturation and Stabilization of Mallory-Denk Bodies
- PMID: 27526095
- PMCID: PMC4985067
- DOI: 10.1371/journal.pone.0161083
p62/Sequestosome-1 Is Indispensable for Maturation and Stabilization of Mallory-Denk Bodies
Abstract
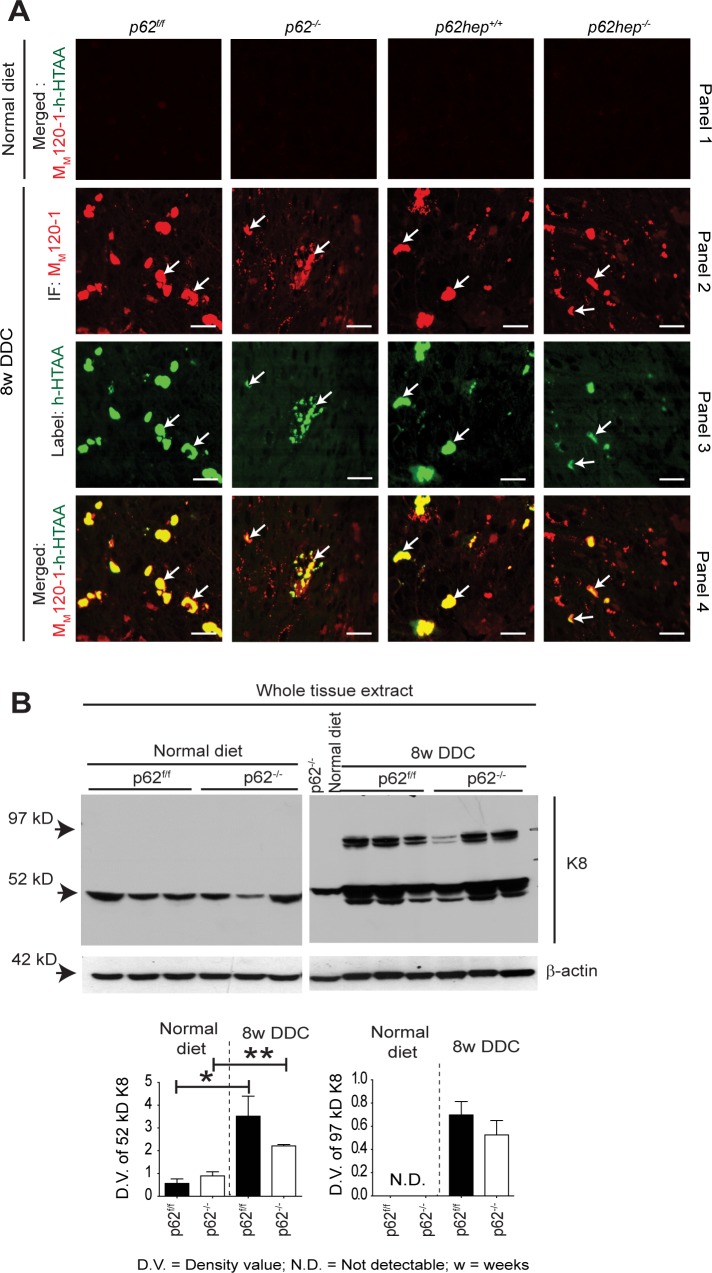
Mallory-Denk bodies (MDBs) are hepatocytic protein aggregates found in steatohepatitis and several other chronic liver diseases as well as hepatocellular carcinoma. MDBs are mainly composed of phosphorylated keratins and stress protein p62/Sequestosome-1 (p62), which is a common component of cytoplasmic aggregates in a variety of protein aggregation diseases. In contrast to the well-established role of keratins, the role of p62 in MDB pathogenesis is still elusive. We have generated total and hepatocyte-specific p62 knockout mice, fed them with 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) to induce MDBs and allowed the mice to recover from DDC intoxication on a standard diet to investigate the role of p62 in MDB formation and elimination. In the absence of p62, smaller, granular and less distinct MDBs appeared, which failed to mature to larger and compact inclusions. Moreover, p62 deficiency impaired the binding of other proteins such as NBR1 and Hsp25 to MDBs and altered the cellular defense mechanism by downregulation of Nrf2 target genes. Upon recovery from DDC intoxication on a standard diet, there was an enhanced reduction of p62-deficient MDBs, which was accompanied by a pronounced decrease in ubiquitinated proteins. Our data provide strong evidence that keratin aggregation is the initial step in MDB formation in steatohepatitis-related mouse models. Interaction of p62 with keratin aggregates then leads to maturation i.e., enlargement and stabilization of the MDBs as well as recruitment of other MDB-associated proteins.
Conflict of interest statement
Figures







Similar articles
-
Oxidative stress, Nrf2 and keratin up-regulation associate with Mallory-Denk body formation in mouse erythropoietic protoporphyria.Hepatology. 2012 Jul;56(1):322-31. doi: 10.1002/hep.25664. Epub 2012 Apr 25. Hepatology. 2012. PMID: 22334478 Free PMC article.
-
Cross β-sheet conformation of keratin 8 is a specific feature of Mallory-Denk bodies compared with other hepatocyte inclusions.Gastroenterology. 2011 Sep;141(3):1080-1090.e1-7. doi: 10.1053/j.gastro.2011.05.039. Epub 2011 May 27. Gastroenterology. 2011. PMID: 21699779
-
The genetic background modulates susceptibility to mouse liver Mallory-Denk body formation and liver injury.Hepatology. 2008 Sep;48(3):943-52. doi: 10.1002/hep.22436. Hepatology. 2008. PMID: 18697208
-
From Mallory to Mallory-Denk bodies: what, how and why?Exp Cell Res. 2007 Jun 10;313(10):2033-49. doi: 10.1016/j.yexcr.2007.04.024. Epub 2007 Apr 27. Exp Cell Res. 2007. PMID: 17531973 Review.
-
Mallory-Denk-bodies: lessons from keratin-containing hepatic inclusion bodies.Biochim Biophys Acta. 2008 Dec;1782(12):764-74. doi: 10.1016/j.bbadis.2008.08.008. Epub 2008 Sep 6. Biochim Biophys Acta. 2008. PMID: 18805482 Review.
Cited by
-
Arsenite exposure suppresses adipogenesis, mitochondrial biogenesis and thermogenesis via autophagy inhibition in brown adipose tissue.Sci Rep. 2019 Oct 8;9(1):14464. doi: 10.1038/s41598-019-50965-9. Sci Rep. 2019. PMID: 31594991 Free PMC article.
-
Target acquired: Selective autophagy in cardiometabolic disease.Sci Signal. 2017 Feb 28;10(468):eaag2298. doi: 10.1126/scisignal.aag2298. Sci Signal. 2017. PMID: 28246200 Free PMC article. Review.
-
NAFLD (MASLD)/NASH (MASH): Does It Bother to Label at All? A Comprehensive Narrative Review.Int J Mol Sci. 2024 Aug 2;25(15):8462. doi: 10.3390/ijms25158462. Int J Mol Sci. 2024. PMID: 39126031 Free PMC article. Review.
-
Utility of Human Relevant Preclinical Animal Models in Navigating NAFLD to MAFLD Paradigm.Int J Mol Sci. 2022 Nov 25;23(23):14762. doi: 10.3390/ijms232314762. Int J Mol Sci. 2022. PMID: 36499091 Free PMC article. Review.
-
Dual roles of p62/SQSTM1 in the injury and recovery phases of acetaminophen-induced liver injury in mice.Acta Pharm Sin B. 2021 Dec;11(12):3791-3805. doi: 10.1016/j.apsb.2021.11.010. Epub 2021 Nov 16. Acta Pharm Sin B. 2021. PMID: 35024307 Free PMC article.
References
-
- Dobson CM. Protein folding and misfolding. Nature. 2003;426(6968):884–90. - PubMed
-
- Reynaud E. Protein Misfolding and Degenerative Diseases. Nature Education. 2010;3(9):28.
-
- Zatloukal K, French SW, Stumptner C, Strnad P, Harada M, Toivola DM, et al. From Mallory to Mallory-Denk bodies: what, how and why? Experimental Cell Research. 2007;313(10):2033–49. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
