

Structural basis of control of inward rectifier Kir2 channel gating by bulk anionic phospholipids
- PMID: 27527100
- PMCID: PMC5004336
- DOI: 10.1085/jgp.201611616
Structural basis of control of inward rectifier Kir2 channel gating by bulk anionic phospholipids
Abstract
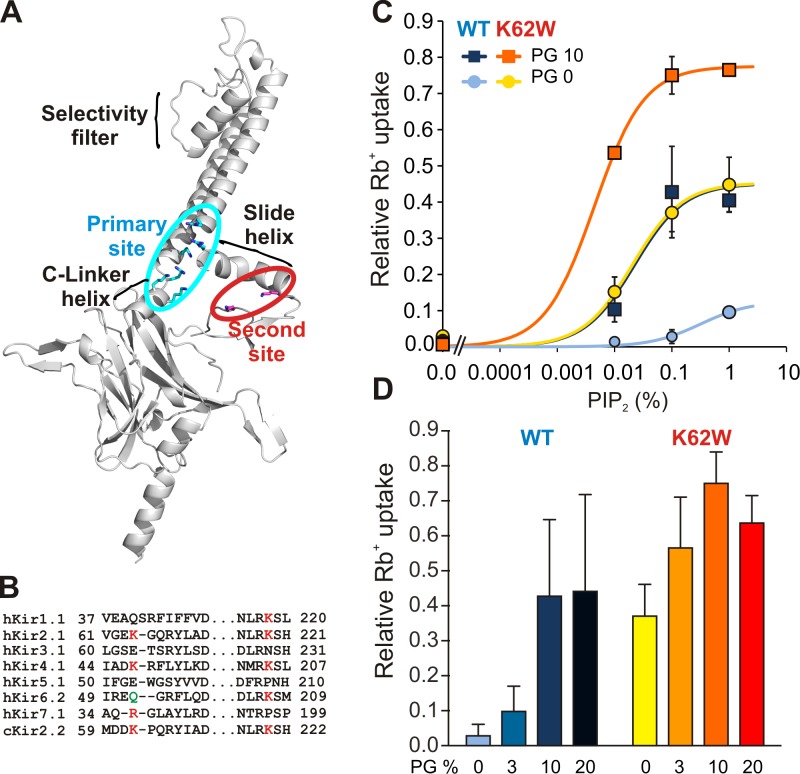
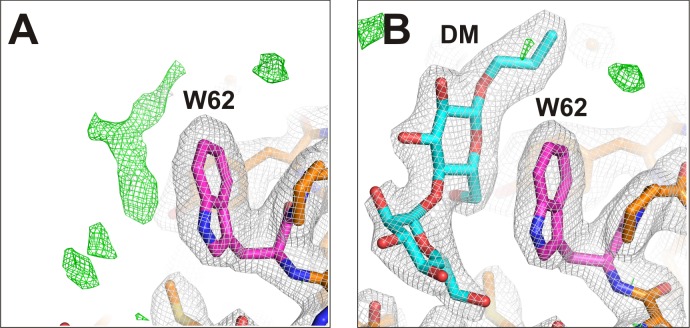
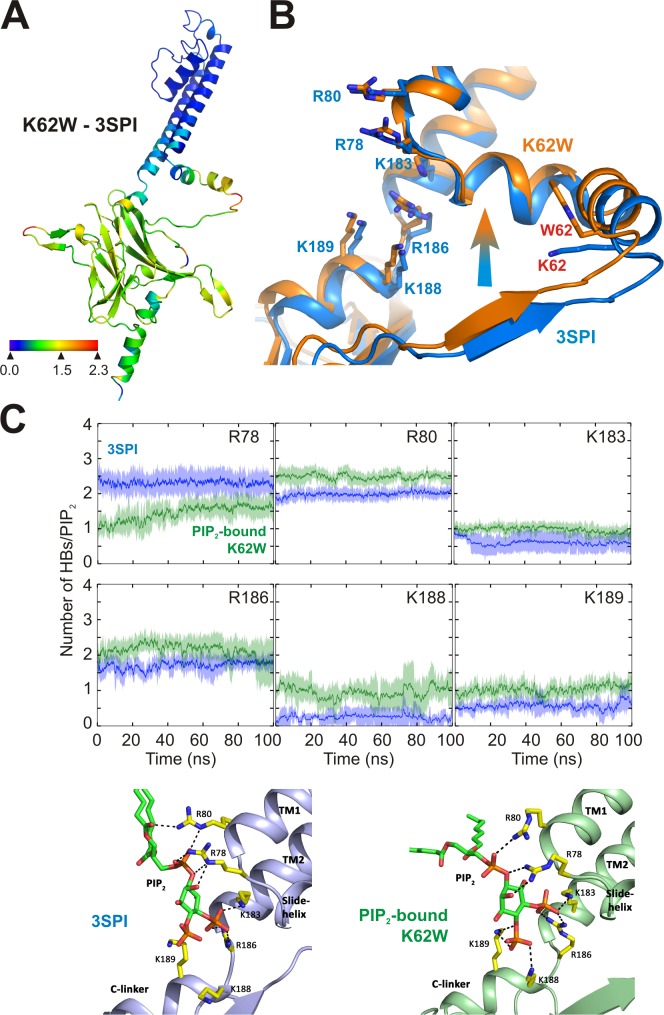
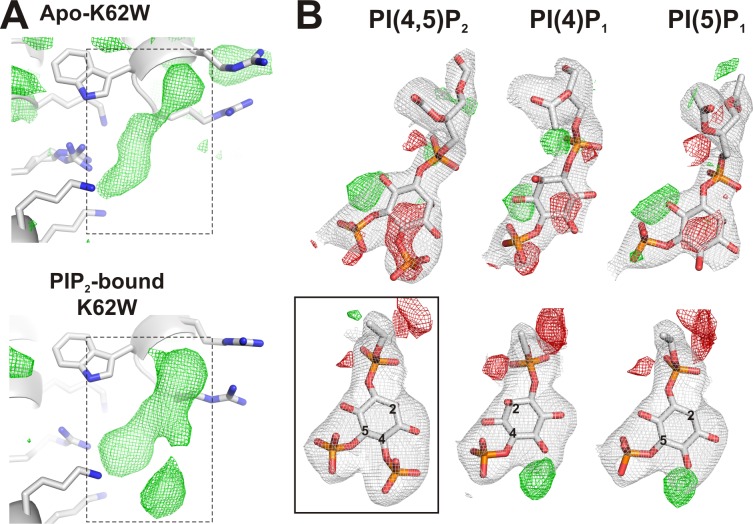
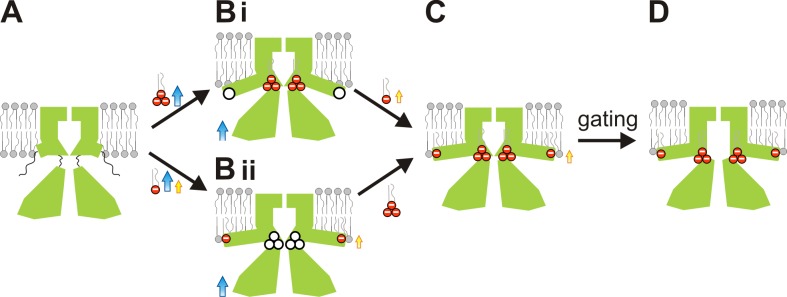
Inward rectifier potassium (Kir) channel activity is controlled by plasma membrane lipids. Phosphatidylinositol-4,5-bisphosphate (PIP2) binding to a primary site is required for opening of classic inward rectifier Kir2.1 and Kir2.2 channels, but interaction of bulk anionic phospholipid (PL(-)) with a distinct second site is required for high PIP2 sensitivity. Here we show that introduction of a lipid-partitioning tryptophan at the second site (K62W) generates high PIP2 sensitivity, even in the absence of PL(-) Furthermore, high-resolution x-ray crystal structures of Kir2.2[K62W], with or without added PIP2 (2.8- and 2.0-Å resolution, respectively), reveal tight tethering of the C-terminal domain (CTD) to the transmembrane domain (TMD) in each condition. Our results suggest a refined model for phospholipid gating in which PL(-) binding at the second site pulls the CTD toward the membrane, inducing the formation of the high-affinity primary PIP2 site and explaining the positive allostery between PL(-) binding and PIP2 sensitivity.
© 2016 Lee et al.
Figures

References
-
- Berendsen H.J.C., Grigera J.R., and Straatsma T.P.. 1987. The missing term in effective pair potentials. J. Phys. Chem. 91:6269–6271. 10.1021/j100308a038 - DOI
-
- Chen V.B., Arendall W.B. III, Headd J.J., Keedy D.A., Immormino R.M., Kapral G.J., Murray L.W., Richardson J.S., and Richardson D.C.. 2010. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66:12–21. 10.1107/S0907444909042073 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
