

A simulation study on the constancy of cardiac energy metabolites during workload transition
- PMID: 27530892
- PMCID: PMC5275773
- DOI: 10.1113/JP272598
A simulation study on the constancy of cardiac energy metabolites during workload transition
Abstract
Key points: The cardiac energy metabolites such as ATP, phosphocreatine, ADP and NADH are kept relatively constant during physiological cardiac workload transition. How this is accomplished is not yet clarified, though Ca2+ has been suggested to be one of the possible mechanisms. We constructed a detailed mathematical model of cardiac mitochondria based on experimental data and studied whether known Ca2+ -dependent regulation mechanisms play roles in the metabolite constancy. Model simulations revealed that the Ca2+ -dependent regulation mechanisms have important roles under the in vitro condition of isolated mitochondria where malate and glutamate were mitochondrial substrates, while they have only a minor role and the composition of substrates has marked influence on the metabolite constancy during workload transition under the simulated in vivo condition where many substrates exist. These results help us understand the regulation mechanisms of cardiac energy metabolism during physiological cardiac workload transition.
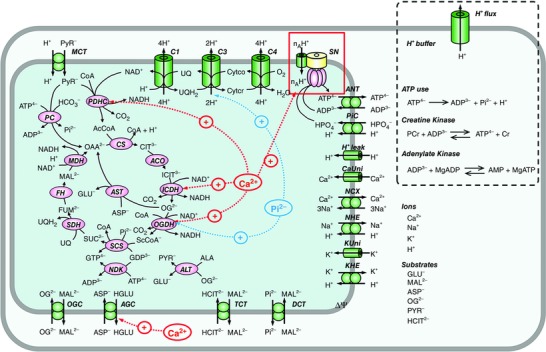
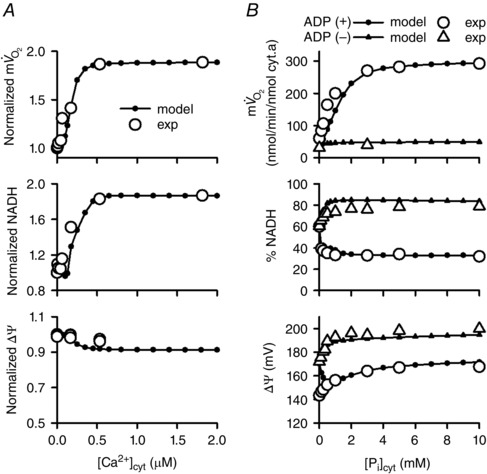
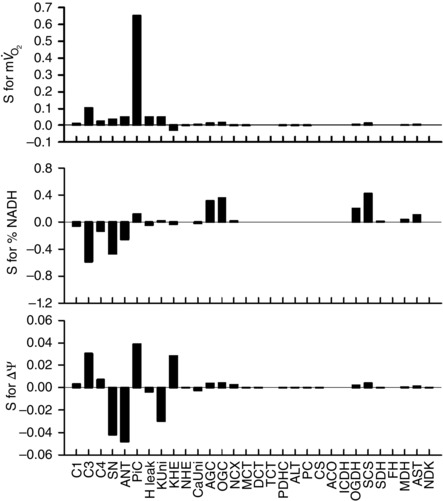
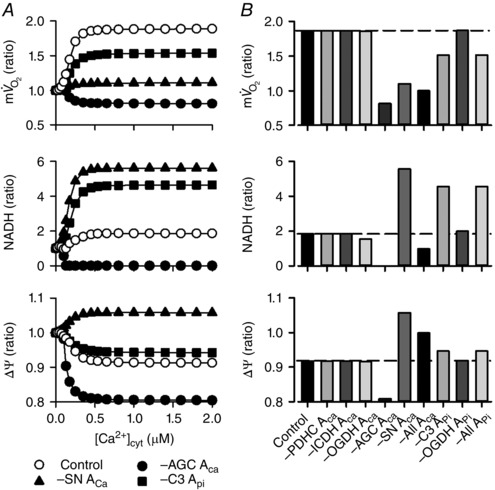
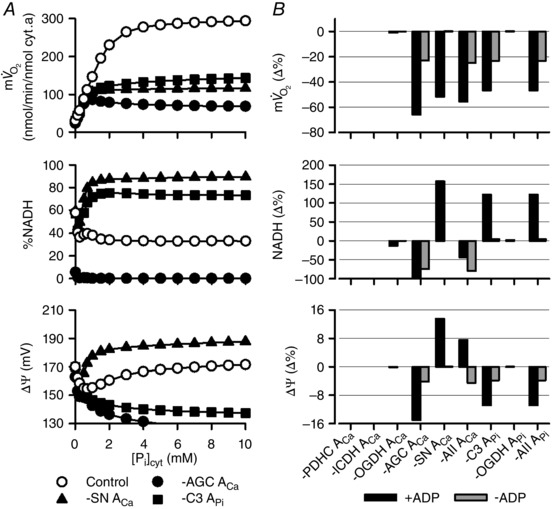
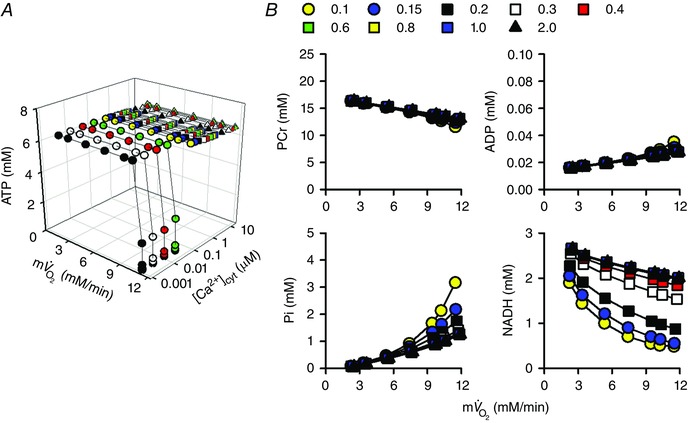
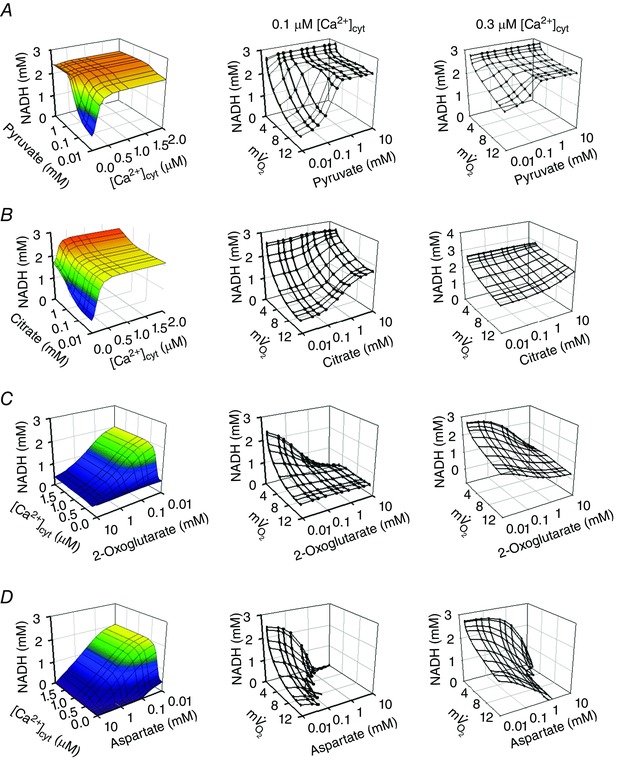
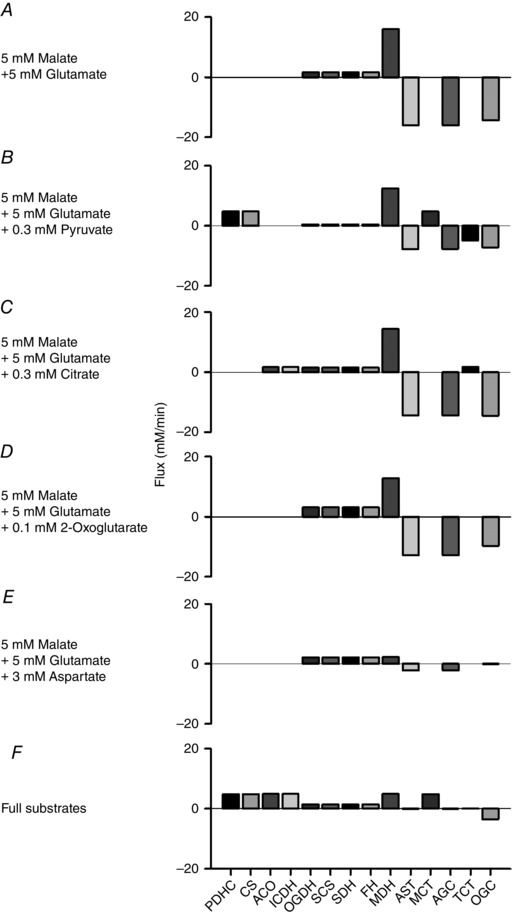
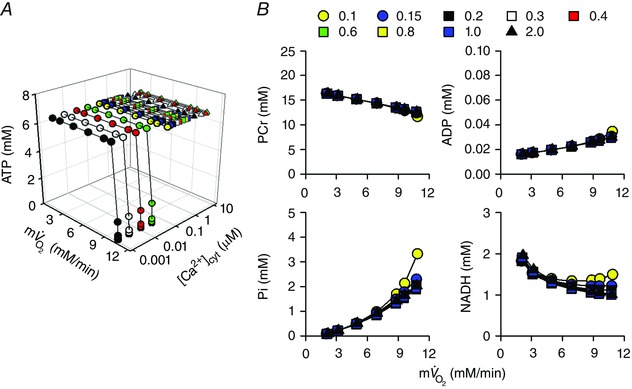
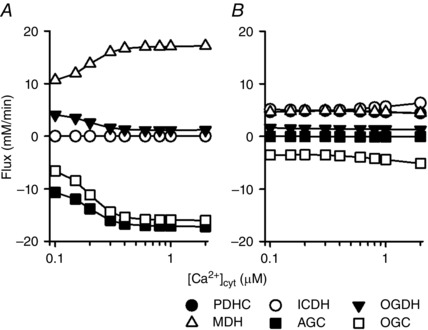
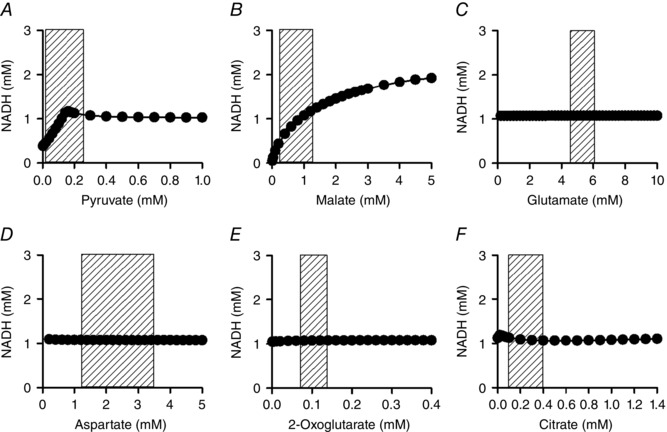
Abstract: The cardiac energy metabolites such as ATP, phosphocreatine, ADP and NADH are kept relatively constant over a wide range of cardiac workload, though the mechanisms are not yet clarified. One possible regulator of mitochondrial metabolism is Ca2+ , because it activates several mitochondrial enzymes and transporters. Here we constructed a mathematical model of cardiac mitochondria, including oxidative phosphorylation, substrate metabolism and ion/substrate transporters, based on experimental data, and studied whether the Ca2+ -dependent activation mechanisms play roles in metabolite constancy. Under the in vitro condition of isolated mitochondria, where malate and glutamate were used as mitochondrial substrates, the model well reproduced the Ca2+ and inorganic phosphate (Pi ) dependences of oxygen consumption, NADH level and mitochondrial membrane potential. The Ca2+ -dependent activations of the aspartate/glutamate carrier and the F1 Fo -ATPase, and the Pi -dependent activation of Complex III were key factors in reproducing the experimental data. When the mitochondrial model was implemented in a simple cardiac cell model, simulation of workload transition revealed that cytoplasmic Ca2+ concentration ([Ca2+ ]cyt ) within the physiological range markedly increased NADH level. However, the addition of pyruvate or citrate attenuated the Ca2+ dependence of NADH during the workload transition. Under the simulated in vivo condition where malate, glutamate, pyruvate, citrate and 2-oxoglutarate were used as mitochondrial substrates, the energy metabolites were more stable during the workload transition and NADH level was almost insensitive to [Ca2+ ]cyt . It was revealed that mitochondrial substrates have a significant influence on metabolite constancy during cardiac workload transition, and Ca2+ has only a minor role under physiological conditions.
Keywords: heart; mathematical modelling; mitochondria.
© 2016 The Authors. The Journal of Physiology © 2016 The Physiological Society.
Figures
References
-
- Albe KR, Butler MH & Wright BE (1990). Cellular concentrations of enzymes and their substrates. J Theor Biol 143, 163–195. - PubMed
-
- Andrienko T, Kuznetsov AV, Kaambre T, Usson Y, Orosco A, Appaix F, Tiivel T, Sikk P, Vendelin M, Margreiter R & Saks VA (2003). Metabolic consequences of functional complexes of mitochondria, myofibrils and sarcoplasmic reticulum in muscle cells. J Exp Biol 206, 2059–2072. - PubMed
-
- Bache RJ, Zhang J, Murakami Y, Zhang Y, Cho YK, Merkle H, Gong G, From AH & Ugurbil K (1999). Myocardial oxygenation at high workstates in hearts with left ventricular hypertrophy. Cardiovasc Res 42, 616–626. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous