

Vpu-Mediated Counteraction of Tetherin Is a Major Determinant of HIV-1 Interferon Resistance
- PMID: 27531907
- PMCID: PMC4992969
- DOI: 10.1128/mBio.00934-16
Vpu-Mediated Counteraction of Tetherin Is a Major Determinant of HIV-1 Interferon Resistance
Abstract
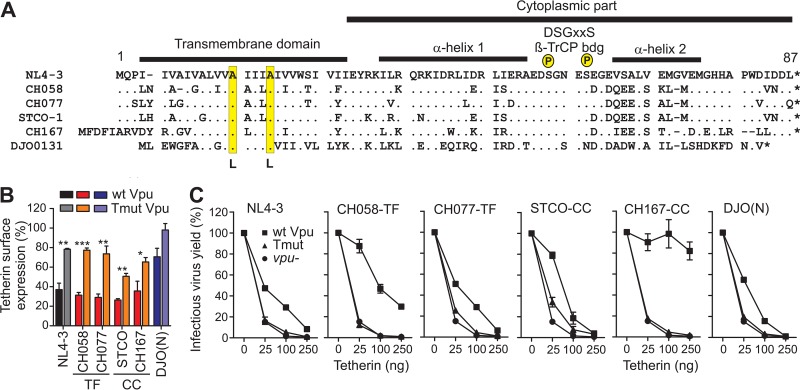
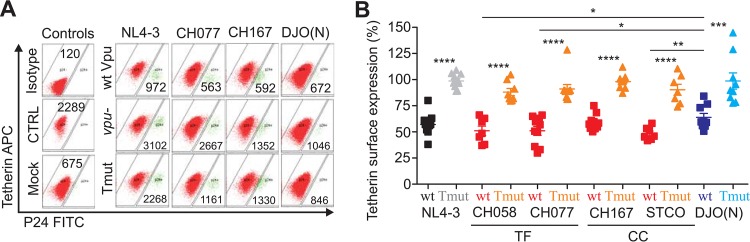
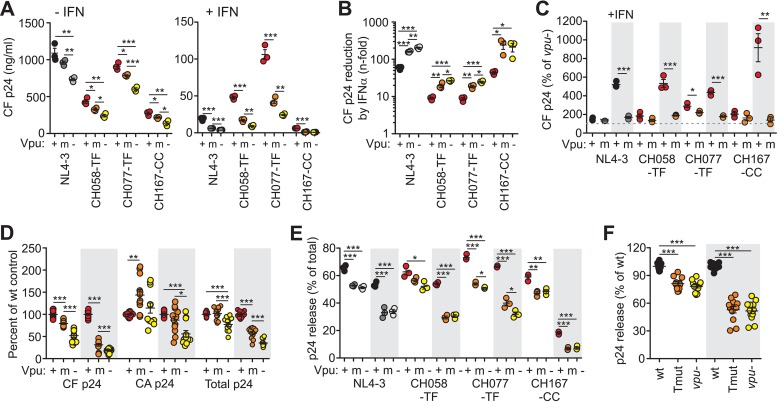
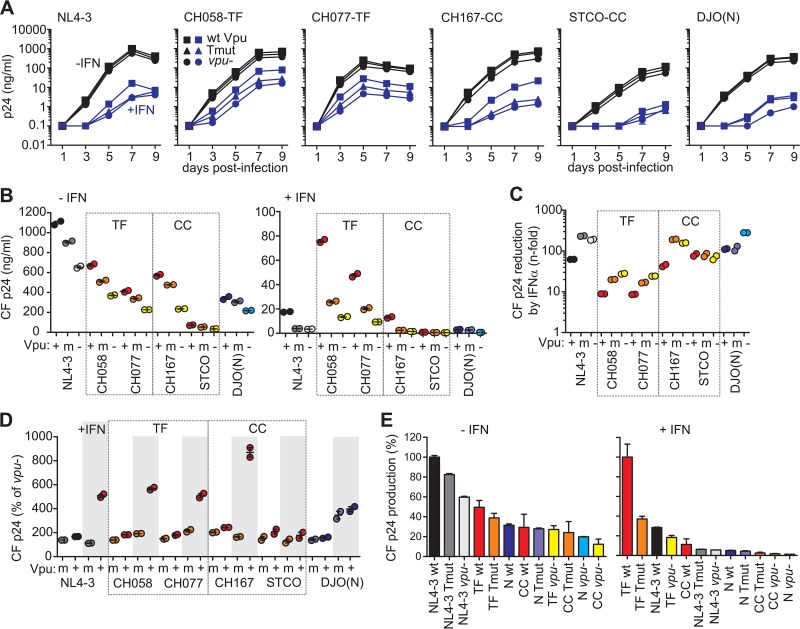
Human immunodeficiency virus type 1 (HIV-1) groups M, N, O, and P are the result of independent zoonotic transmissions of simian immunodeficiency viruses (SIVs) infecting great apes in Africa. Among these, only Vpu proteins of pandemic HIV-1 group M strains evolved potent activity against the restriction factor tetherin, which inhibits virus release from infected cells. Thus, effective Vpu-mediated tetherin antagonism may have been a prerequisite for the global spread of HIV-1. To determine whether this particular function enhances primary HIV-1 replication and interferon resistance, we introduced mutations into the vpu genes of HIV-1 group M and N strains to specifically disrupt their ability to antagonize tetherin, but not other Vpu functions, such as degradation of CD4, down-modulation of CD1d and NTB-A, and suppression of NF-κB activity. Lack of particular human-specific adaptations reduced the ability of HIV-1 group M Vpu proteins to enhance virus production and release from primary CD4(+) T cells at high levels of type I interferon (IFN) from about 5-fold to 2-fold. Interestingly, transmitted founder HIV-1 strains exhibited higher virion release capacity than chronic control HIV-1 strains irrespective of Vpu function, and group M viruses produced higher levels of cell-free virions than an N group HIV-1 strain. Thus, efficient virus release from infected cells seems to play an important role in the spread of HIV-1 in the human population and requires a fully functional Vpu protein that counteracts human tetherin.
Importance: Understanding which human-specific adaptations allowed HIV-1 to cause the AIDS pandemic is of great importance. One feature that distinguishes pandemic HIV-1 group M strains from nonpandemic or rare group O, N, and P viruses is the acquisition of mutations in the accessory Vpu protein that confer potent activity against human tetherin. Adaptation was required because human tetherin has a deletion that renders it resistant to the Nef protein used by the SIV precursor of HIV-1 to antagonize this antiviral factor. It has been suggested that these adaptations in Vpu were critical for the effective spread of HIV-1 M strains, but direct evidence has been lacking. Here, we show that these changes in Vpu significantly enhance virus replication and release in human CD4(+) T cells, particularly in the presence of IFN, thus supporting an important role in the spread of pandemic HIV-1.
Copyright © 2016 Kmiec et al.
Figures
Similar articles
-
Efficient Vpu-Mediated Tetherin Antagonism by an HIV-1 Group O Strain.J Virol. 2017 Feb 28;91(6):e02177-16. doi: 10.1128/JVI.02177-16. Print 2017 Mar 15. J Virol. 2017. PMID: 28077643 Free PMC article.
-
HIV-1 Group P is unable to antagonize human tetherin by Vpu, Env or Nef.Retrovirology. 2011 Dec 15;8:103. doi: 10.1186/1742-4690-8-103. Retrovirology. 2011. PMID: 22171785 Free PMC article.
-
The transmembrane domain of HIV-1 Vpu is sufficient to confer anti-tetherin activity to SIVcpz and SIVgor Vpu proteins: cytoplasmic determinants of Vpu function.Retrovirology. 2013 Mar 20;10:32. doi: 10.1186/1742-4690-10-32. Retrovirology. 2013. PMID: 23514615 Free PMC article.
-
Emerging role of the host restriction factor tetherin in viral immune sensing.J Mol Biol. 2013 Dec 13;425(24):4956-64. doi: 10.1016/j.jmb.2013.09.029. Epub 2013 Sep 26. J Mol Biol. 2013. PMID: 24075872 Review.
-
Modulation of HIV-1-host interaction: role of the Vpu accessory protein.Retrovirology. 2010 Dec 22;7:114. doi: 10.1186/1742-4690-7-114. Retrovirology. 2010. PMID: 21176220 Free PMC article. Review.
Cited by
-
HIV-1 Vpu is a potent transcriptional suppressor of NF-κB-elicited antiviral immune responses.Elife. 2019 Feb 5;8:e41930. doi: 10.7554/eLife.41930. Elife. 2019. PMID: 30717826 Free PMC article.
-
BST-2 promotes survival in circulation and pulmonary metastatic seeding of breast cancer cells.Sci Rep. 2018 Dec 4;8(1):17608. doi: 10.1038/s41598-018-35710-y. Sci Rep. 2018. PMID: 30514852 Free PMC article.
-
Manipulation of Mononuclear Phagocytes by HIV: Implications for Early Transmission Events.Front Immunol. 2019 Sep 24;10:2263. doi: 10.3389/fimmu.2019.02263. eCollection 2019. Front Immunol. 2019. PMID: 31616434 Free PMC article. Review.
-
The HIV-1 transmission bottleneck.Retrovirology. 2017 Mar 23;14(1):22. doi: 10.1186/s12977-017-0343-8. Retrovirology. 2017. PMID: 28335782 Free PMC article.
-
Antiviral factors and their counteraction by HIV-1: many uncovered and more to be discovered.J Mol Cell Biol. 2024 Jul 29;16(2):mjae005. doi: 10.1093/jmcb/mjae005. J Mol Cell Biol. 2024. PMID: 38318650 Free PMC article. Review.
References
-
- Sauter D, Schindler M, Specht A, Landford WN, Münch J, Kim KA, Votteler J, Schubert U, Bibollet-Ruche F, Keele BF, Takehisa J, Ogando Y, Ochsenbauer C, Kappes JC, Ayouba A, Peeters M, Learn GH, Shaw G, Sharp PM, Bieniasz P, Hahn BH, Hatziioannou T, Kirchhoff F. 2009. Tetherin-driven adaptation of Vpu and Nef function and the evolution of pandemic and nonpandemic HIV-1 strains. Cell Host Microbe 6:409–421. doi:10.1016/j.chom.2009.10.004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
