

Human leukocyte Antigen-DM polymorphisms in autoimmune diseases
- PMID: 27534821
- PMCID: PMC5008016
- DOI: 10.1098/rsob.160165
Human leukocyte Antigen-DM polymorphisms in autoimmune diseases
Abstract
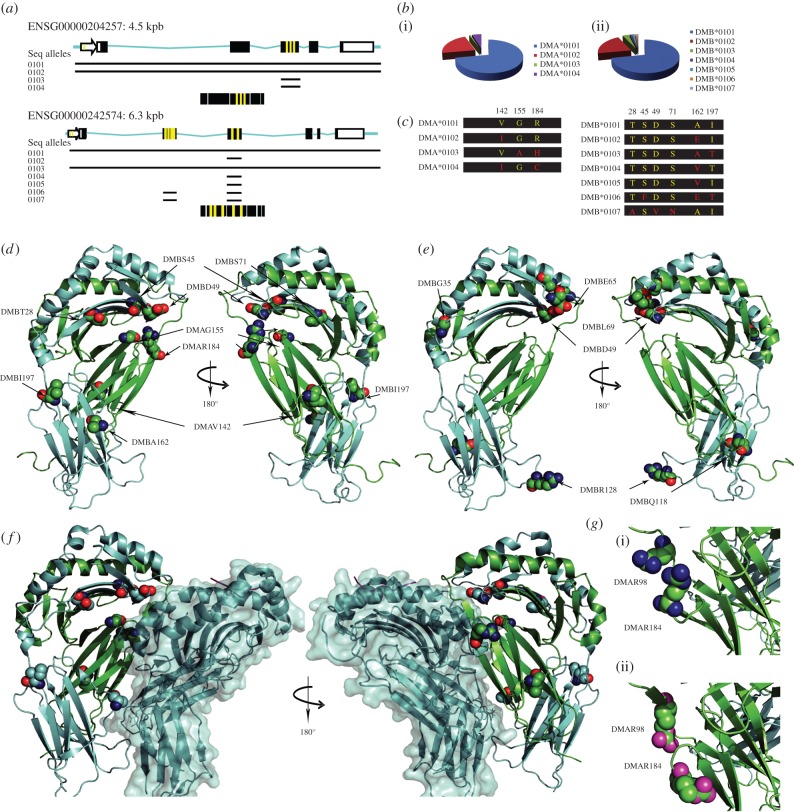
Classical MHC class II (MHCII) proteins present peptides for CD4(+) T-cell surveillance and are by far the most prominent risk factor for a number of autoimmune disorders. To date, many studies have shown that this link between particular MHCII alleles and disease depends on the MHCII's particular ability to bind and present certain peptides in specific physiological contexts. However, less attention has been paid to the non-classical MHCII molecule human leucocyte antigen-DM, which catalyses peptide exchange on classical MHCII proteins acting as a peptide editor. DM function impacts the presentation of both antigenic peptides in the periphery and key self-peptides during T-cell development in the thymus. In this way, DM activity directly influences the response to pathogens, as well as mechanisms of self-tolerance acquisition. While decreased DM editing of particular MHCII proteins has been proposed to be related to autoimmune disorders, no experimental evidence for different DM catalytic properties had been reported until recently. Biochemical and structural investigations, together with new animal models of loss of DM activity, have provided an attractive foundation for identifying different catalytic efficiencies for DM allotypes. Here, we revisit the current knowledge of DM function and discuss how DM function may impart autoimmunity at the organism level.
Keywords: antigen presentation; autoimmunity; human leucocyte antigen-DM; major histocompatibility complex of class II; peptidome; polymorphism.
© 2016 The Authors.
Figures



References
-
- Rai E, Wakeland EK. 2011. Genetic predisposition to autoimmunity: what have we learned? Semin. Immunol. 23, 67–83. (doi:10.1016/j.smim.2011.01.015) - DOI - PubMed
-
- Wucherpfennig KW, Sethi D. 2011. T cell receptor recognition of self and foreign antigens in the induction of autoimmunity. Semin. Immunol. 23, 84–91. (doi:10.1016/j.smim.2011.01.007) - DOI - PMC - PubMed
-
- Shiina T, Hosomochi K, Kulski JK. 2009. The HLA genomic loci map: expression, interaction, diversity and disease. J. Hum. Genet. 54, 15–39. (doi:10.1038/jhg.2008.5) - DOI - PubMed
-
- Schlosstein L, Terasaki PI, Bluestone R, Pearson CM. 1973. High association of an HL-A antigen, W27, with ankylosing spondylitis. N. Engl. J. Med. 288, 704–706. (doi:10.1056/NEJM197304052881403) - DOI - PubMed
-
- Fernando MM, Stevens CR, Walsh EC, De Jager PL, Goyette P, Plenge RM, Vyse TJ, Rioux JD. 2008. Defining the role of the MHC in autoimmunity: a review and pooled analysis. PLoS Genet. 4, e1000024 (doi:10.1371/journal.pgen.1000024) - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials