

HIV and Cocaine Impact Glial Metabolism: Energy Sensor AMP-activated protein kinase Role in Mitochondrial Biogenesis and Epigenetic Remodeling
- PMID: 27535703
- PMCID: PMC4989157
- DOI: 10.1038/srep31784
HIV and Cocaine Impact Glial Metabolism: Energy Sensor AMP-activated protein kinase Role in Mitochondrial Biogenesis and Epigenetic Remodeling
Abstract
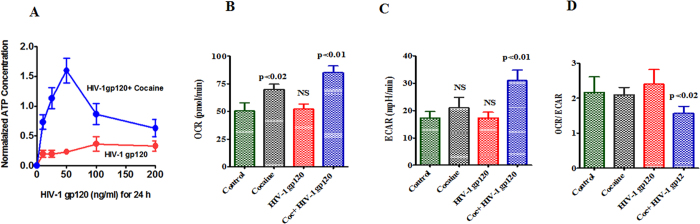
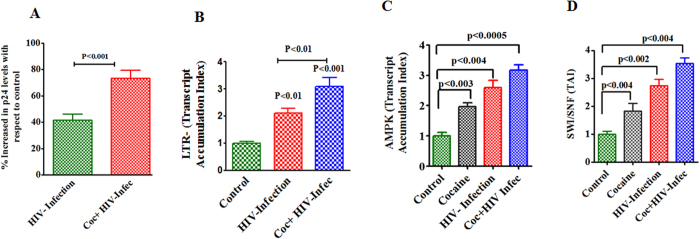
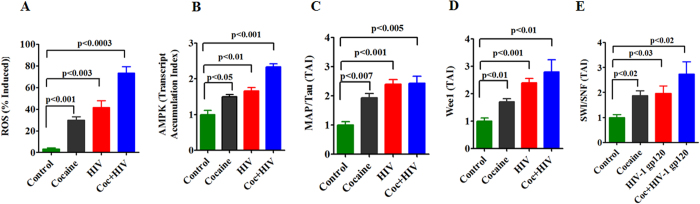
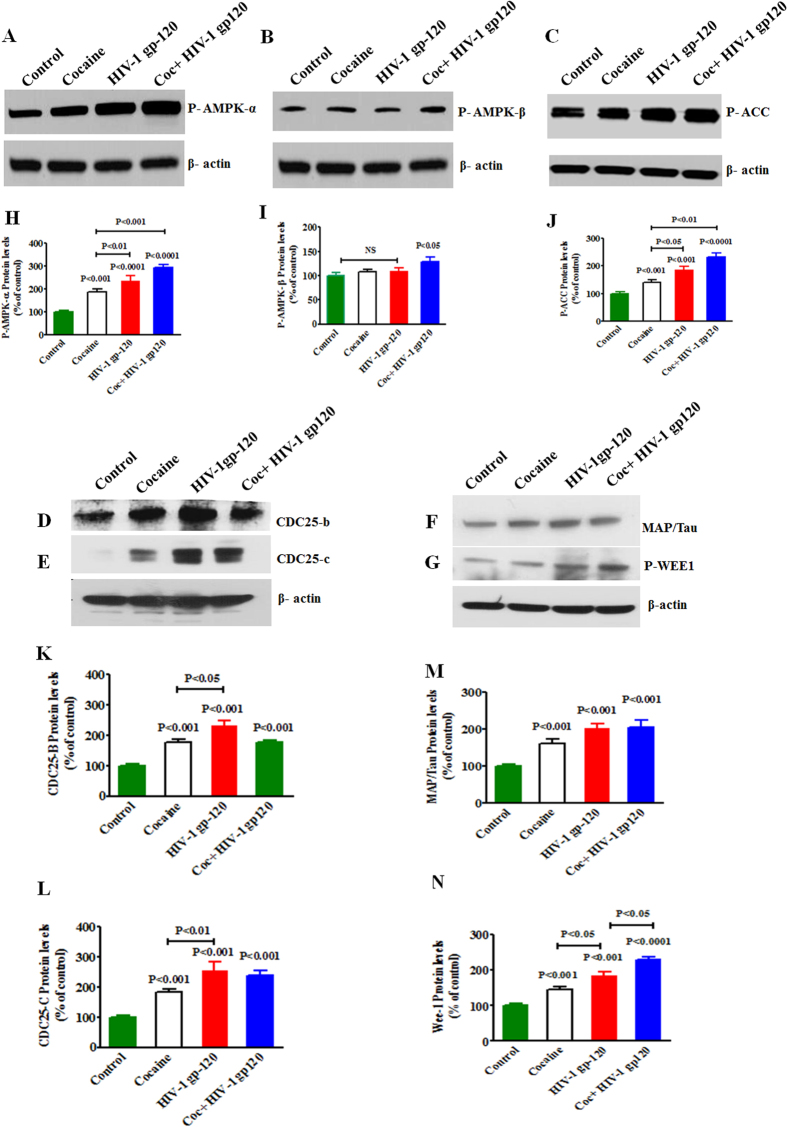
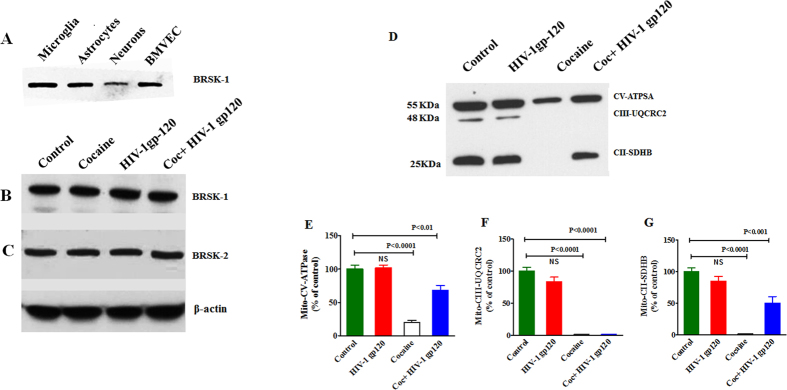
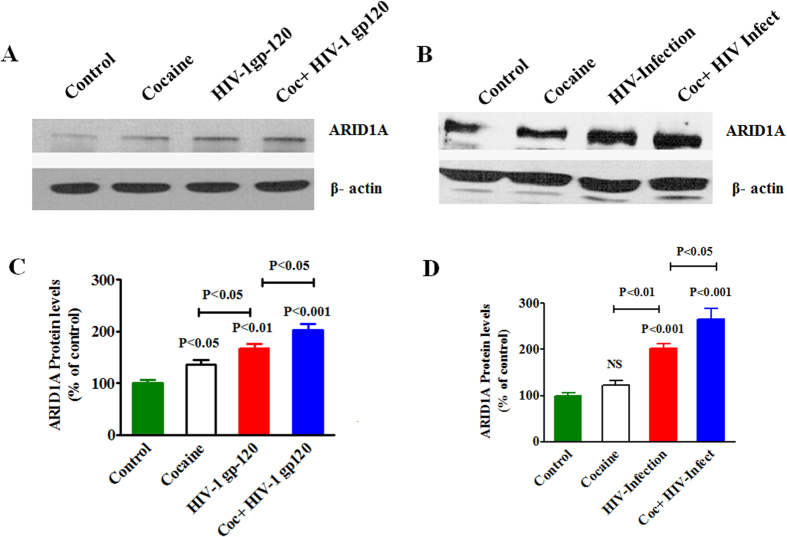
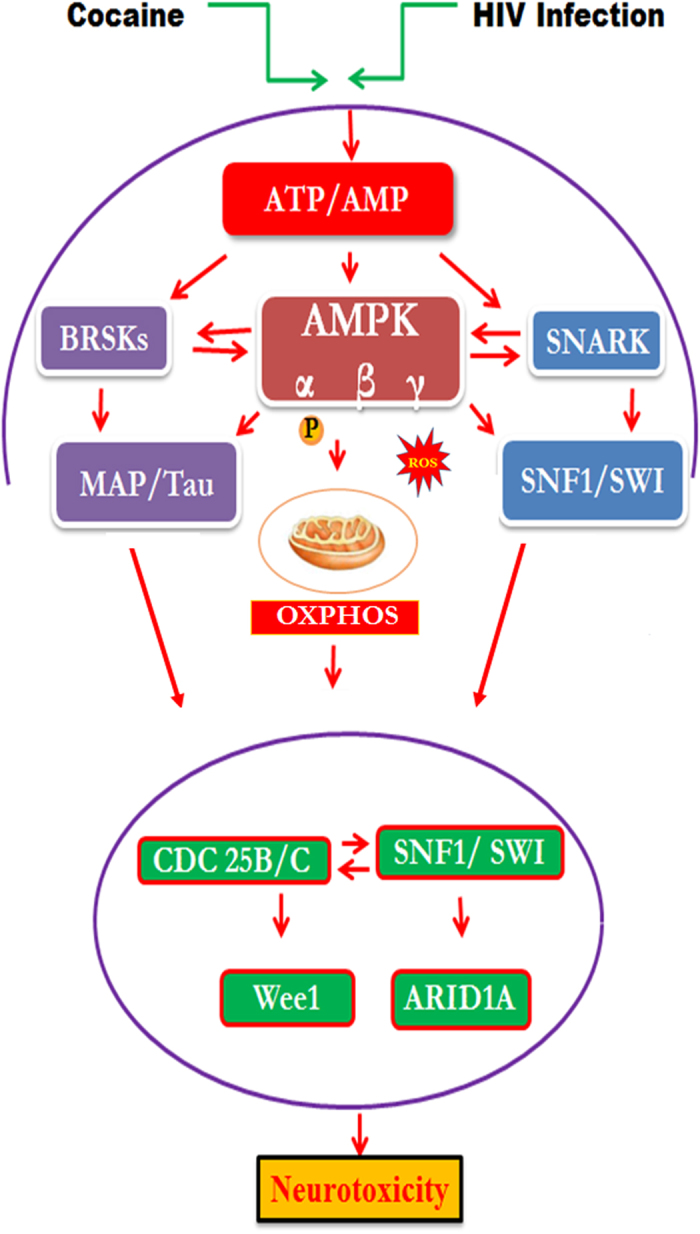
HIV infection and cocaine use have been identified as risk factors for triggering neuronal dysfunction. In the central nervous system (CNS), energy resource and metabolic function are regulated by astroglia. Glia is the major reservoir of HIV infection and disease progression in CNS. However, the role of cocaine in accelerating HIV associated energy deficit and its impact on neuronal dysfunction has not been elucidated yet. The aim of this study is to elucidate the molecular mechanism of HIV associated neuropathogenesis in cocaine abuse and how it accelerates the energy sensor AMPKs and its subsequent effect on mitochondrial oxidative phosphorylation (OXPHOS), BRSKs, CDC25B/C, MAP/Tau, Wee1 and epigenetics remodeling complex SWI/SNF. Results showed that cocaine exposure during HIV infection significantly increased the level of p24, reactive oxygen species (ROS), ATP-utilization and upregulated energy sensor AMPKs, CDC25B/C, MAP/Tau and Wee1 protein expression. Increased ROS production subsequently inhibits OCR/ECAR ratio and OXPHOS, and eventually upregulate epigenetics remodeling complex SWI/SNF in CHME-5 cells. These results suggest that HIV infection induced energy deficit and metabolic dysfunction is accelerated by cocaine inducing energy sensor AMPKs, mitochondrial biogenesis and chromatin remodeling complex SWI/SNF activation, which may lead to neuroAIDS disease progression.
Figures
Similar articles
-
The effects of cocaine on HIV transcription.J Neurovirol. 2016 Jun;22(3):261-74. doi: 10.1007/s13365-015-0398-z. Epub 2015 Nov 16. J Neurovirol. 2016. PMID: 26572787 Free PMC article. Review.
-
The role of mitochondria in cocaine addiction.Biochem J. 2021 Feb 26;478(4):749-764. doi: 10.1042/BCJ20200615. Biochem J. 2021. PMID: 33626141 Review.
-
HIV-1 Tat and cocaine impact astrocytic energy reservoir influence on miRNA epigenetic regulation.Genomics. 2021 Nov;113(6):3461-3475. doi: 10.1016/j.ygeno.2021.08.013. Epub 2021 Aug 19. Genomics. 2021. PMID: 34418497 Free PMC article.
-
Mitochondrial dysfunction and caspase activation in rat cortical neurons treated with cocaine or amphetamine.Brain Res. 2006 May 17;1089(1):44-54. doi: 10.1016/j.brainres.2006.03.061. Epub 2006 Apr 25. Brain Res. 2006. PMID: 16638611
-
HIV-1 neuropathogenesis: glial mechanisms revealed through substance abuse.J Neurochem. 2007 Feb;100(3):567-86. doi: 10.1111/j.1471-4159.2006.04227.x. Epub 2006 Dec 1. J Neurochem. 2007. PMID: 17173547 Free PMC article. Review.
Cited by
-
HIV-1 Tat and cocaine impact astrocytic energy reservoirs and epigenetic regulation by influencing the LINC01133-hsa-miR-4726-5p-NDUFA9 axis.Mol Ther Nucleic Acids. 2022 Jul 6;29:243-258. doi: 10.1016/j.omtn.2022.07.001. eCollection 2022 Sep 13. Mol Ther Nucleic Acids. 2022. PMID: 35892093 Free PMC article.
-
Cocaine Differentially Affects Mitochondrial Function Depending on Exposure Time.Int J Mol Sci. 2025 Feb 27;26(5):2131. doi: 10.3390/ijms26052131. Int J Mol Sci. 2025. PMID: 40076756 Free PMC article.
-
Human microglial models to study HIV infection and neuropathogenesis: a literature overview and comparative analyses.J Neurovirol. 2022 Feb;28(1):64-91. doi: 10.1007/s13365-021-01049-w. Epub 2022 Feb 9. J Neurovirol. 2022. PMID: 35138593 Free PMC article. Review.
-
Suppression of HIV and cocaine-induced neurotoxicity and inflammation by cell penetrable itaconate esters.bioRxiv [Preprint]. 2023 Sep 26:2023.09.25.559154. doi: 10.1101/2023.09.25.559154. bioRxiv. 2023. Update in: J Neurovirol. 2024 Aug;30(4):337-352. doi: 10.1007/s13365-024-01216-9. PMID: 37808776 Free PMC article. Updated. Preprint.
-
Suppression of HIV-TAT and cocaine-induced neurotoxicity and inflammation by cell penetrable itaconate esters.J Neurovirol. 2024 Aug;30(4):337-352. doi: 10.1007/s13365-024-01216-9. Epub 2024 Jun 17. J Neurovirol. 2024. PMID: 38884890 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
