

Salt Induces Features of a Dormancy-Like State in Seeds of Eutrema (Thellungiella) salsugineum, a Halophytic Relative of Arabidopsis
- PMID: 27536302
- PMCID: PMC4971027
- DOI: 10.3389/fpls.2016.01071
Salt Induces Features of a Dormancy-Like State in Seeds of Eutrema (Thellungiella) salsugineum, a Halophytic Relative of Arabidopsis
Abstract
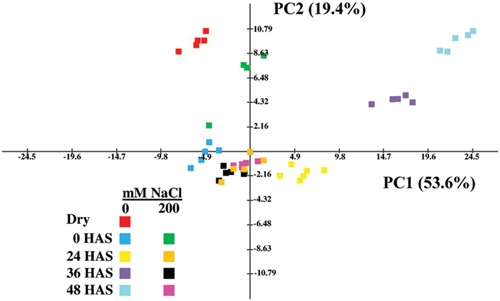
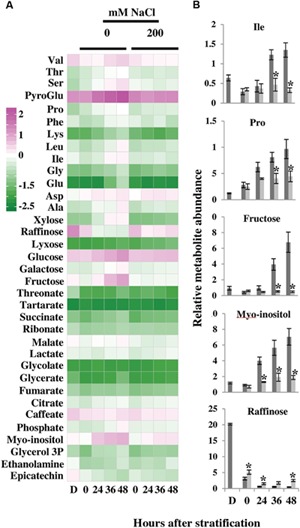
The salinization of land is a major factor limiting crop production worldwide. Halophytes adapted to high levels of salinity are likely to possess useful genes for improving crop tolerance to salt stress. In addition, halophytes could provide a food source on marginal lands. However, despite halophytes being salt-tolerant plants, the seeds of several halophytic species will not germinate on saline soils. Yet, little is understood regarding biochemical and gene expression changes underlying salt-mediated inhibition of halophyte seed germination. We have used the halophytic Arabidopsis relative model system, Eutrema (Thellungiella) salsugineum to explore salt-mediated inhibition of germination. We show that E. salsugineum seed germination is inhibited by salt to a far greater extent than in Arabidopsis, and that this inhibition is in response to the osmotic component of salt exposure. E. salsugineum seeds remain viable even when germination is completely inhibited, and germination resumes once seeds are transferred to non-saline conditions. Moreover, removal of the seed coat from salt-treated seeds allows embryos to germinate on salt-containing medium. Mobilization of seed storage reserves is restricted in salt-treated seeds, while many germination-associated metabolic changes are arrested or progress to a lower extent. Salt-exposed seeds are further characterized by a reduced GA/ABA ratio and increased expression of the germination repressor genes, RGL2, ABI5, and DOG1. Furthermore, a salt-mediated increase in expression of a LATE EMBRYOGENESIS ABUNDANT gene and accretion of metabolites involved in osmoprotection indicates induction of processes associated with stress tolerance, and accumulation of easily mobilized carbon reserves. Overall, our results suggest that salt inhibits E. salsugineum seed germination by inducing a seed state with molecular features of dormancy while a physical constraint to radicle emergence is provided by the seed coat layers. This seed state could facilitate survival on saline soils until a rain event(s) increases soil water potential indicating favorable conditions for seed germination and establishment of salt-tolerant E. salsugineum seedlings.
Keywords: Arabidopsis relative; Brassicaceae; Eutrema salsugineum; dormancy; extremophile plants; halophyte; salt stress; seed germination.
Figures







Similar articles
-
Halophytism: What Have We Learnt From Arabidopsis thaliana Relative Model Systems?Plant Physiol. 2018 Nov;178(3):972-988. doi: 10.1104/pp.18.00863. Epub 2018 Sep 20. Plant Physiol. 2018. PMID: 30237204 Free PMC article. Review.
-
Comparative study of the aldehyde dehydrogenase (ALDH) gene superfamily in the glycophyte Arabidopsis thaliana and Eutrema halophytes.Ann Bot. 2015 Feb;115(3):465-79. doi: 10.1093/aob/mcu152. Epub 2014 Aug 1. Ann Bot. 2015. PMID: 25085467 Free PMC article.
-
Anastatica hierochuntica, an Arabidopsis Desert Relative, Is Tolerant to Multiple Abiotic Stresses and Exhibits Species-Specific and Common Stress Tolerance Strategies with Its Halophytic Relative, Eutrema (Thellungiella) salsugineum.Front Plant Sci. 2017 Jan 17;7:1992. doi: 10.3389/fpls.2016.01992. eCollection 2016. Front Plant Sci. 2017. PMID: 28144244 Free PMC article.
-
Growth platform-dependent and -independent phenotypic and metabolic responses of Arabidopsis and its halophytic relative, Eutrema salsugineum, to salt stress.Plant Physiol. 2013 Jul;162(3):1583-98. doi: 10.1104/pp.113.217844. Epub 2013 Jun 4. Plant Physiol. 2013. PMID: 23735509 Free PMC article.
-
Adaptative Mechanisms of Halophytic Eutrema salsugineum Encountering Saline Environment.Front Plant Sci. 2022 Jun 28;13:909527. doi: 10.3389/fpls.2022.909527. eCollection 2022. Front Plant Sci. 2022. PMID: 35837468 Free PMC article. Review.
Cited by
-
Predicting and testing a gene network regulating seed germination in Arabidopsis.PeerJ. 2025 Jul 7;13:e19599. doi: 10.7717/peerj.19599. eCollection 2025. PeerJ. 2025. PMID: 40642330 Free PMC article.
-
Unraveling the role of urea hydrolysis in salt stress response during seed germination and seedling growth in Arabidopsis thaliana.Elife. 2024 Jul 22;13:e96797. doi: 10.7554/eLife.96797. Elife. 2024. PMID: 39037769 Free PMC article.
-
Halophytism: What Have We Learnt From Arabidopsis thaliana Relative Model Systems?Plant Physiol. 2018 Nov;178(3):972-988. doi: 10.1104/pp.18.00863. Epub 2018 Sep 20. Plant Physiol. 2018. PMID: 30237204 Free PMC article. Review.
-
Negative regulation of seed germination by maternal AFB1 and AFB5 in Arabidopsis.Biosci Rep. 2022 Sep 30;42(9):BSR20221504. doi: 10.1042/BSR20221504. Biosci Rep. 2022. PMID: 36039862 Free PMC article.
-
ERECTA receptor-kinases play a key role in the appropriate timing of seed germination under changing salinity.J Exp Bot. 2019 Nov 18;70(21):6417-6435. doi: 10.1093/jxb/erz385. J Exp Bot. 2019. PMID: 31504732 Free PMC article.
References
-
- Al-Hawija B. N., Partzsch M., Hensen I. (2012). Effects of temperature, salinity and cold stratification on seed germination in halophytes. Nord. J. Bot. 30 627–634. 10.1111/j.1756-1051.2012.01314.x - DOI
-
- Baskin C. C., Baskin J. M. (1998). Seeds; Ecology, Biogeography and Evolution of Dormancy and Germination. San Diego, CA: Academic Press.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases