

Identification and Ultrastructural Characterization of a Novel Nuclear Degradation Complex in Differentiating Lens Fiber Cells
- PMID: 27536868
- PMCID: PMC4990417
- DOI: 10.1371/journal.pone.0160785
Identification and Ultrastructural Characterization of a Novel Nuclear Degradation Complex in Differentiating Lens Fiber Cells
Abstract
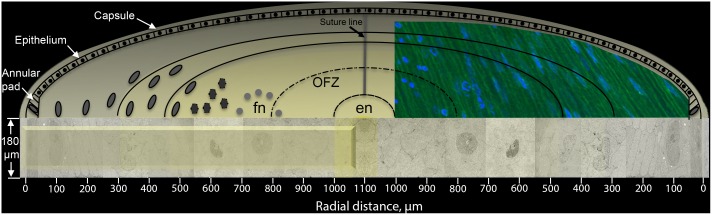
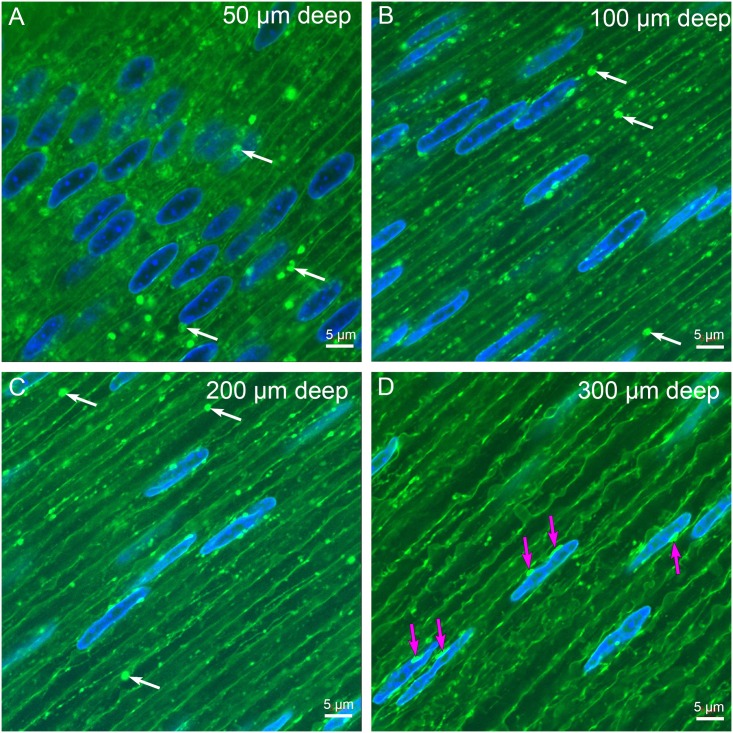
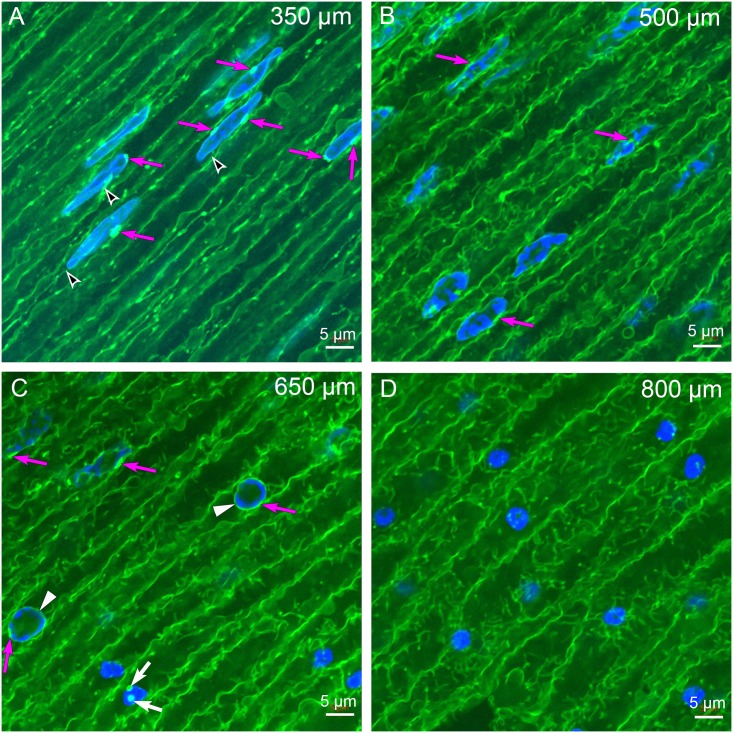
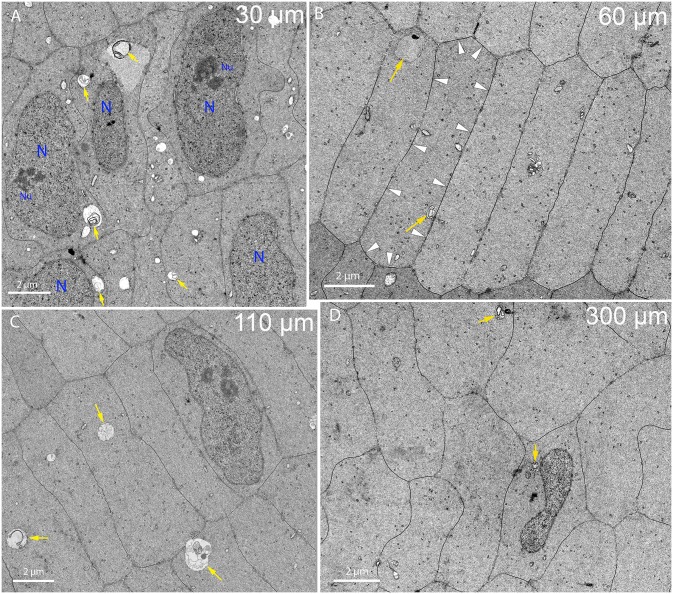
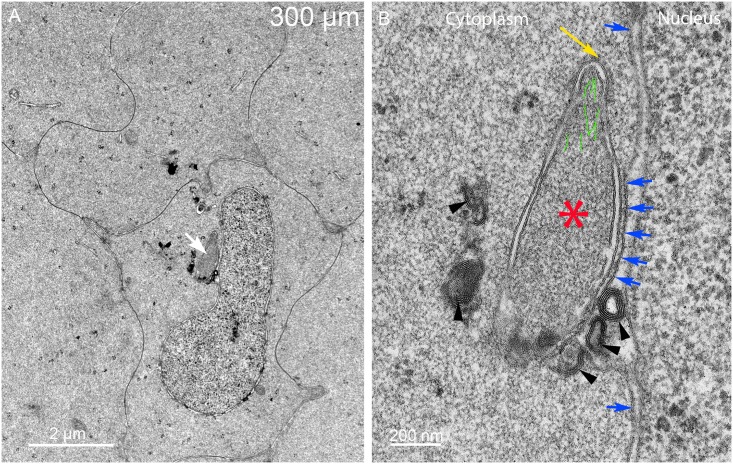
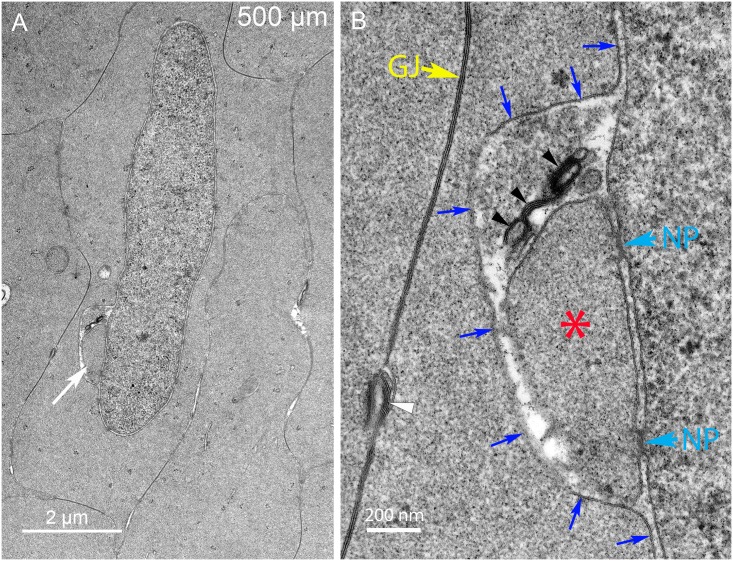
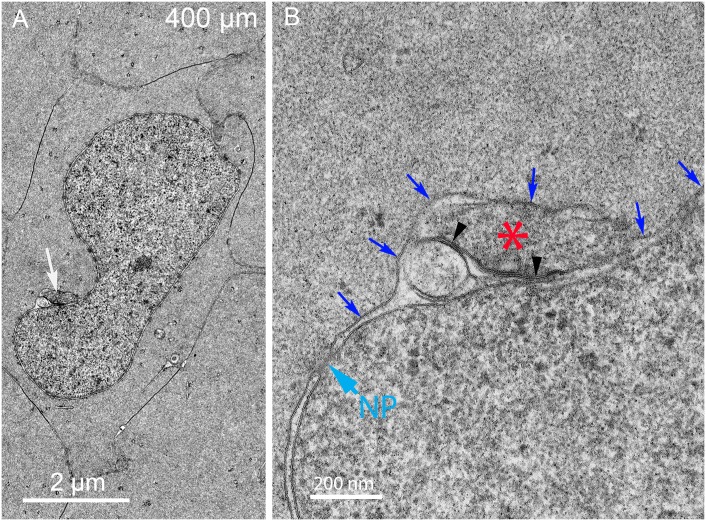
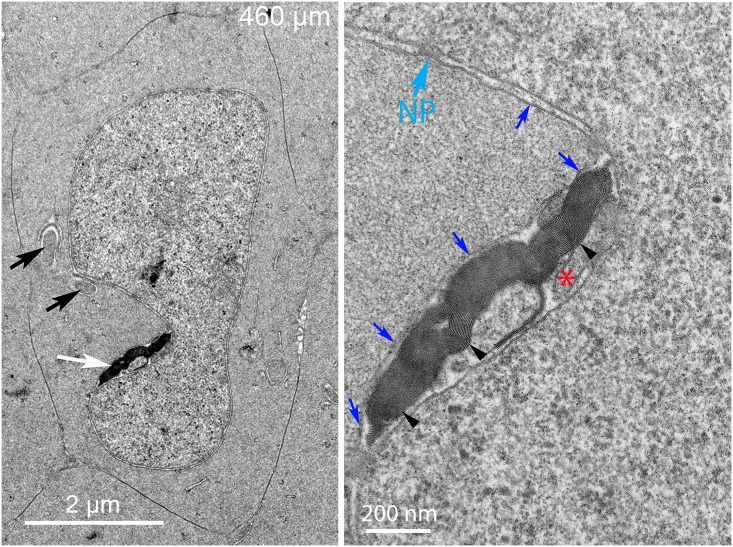
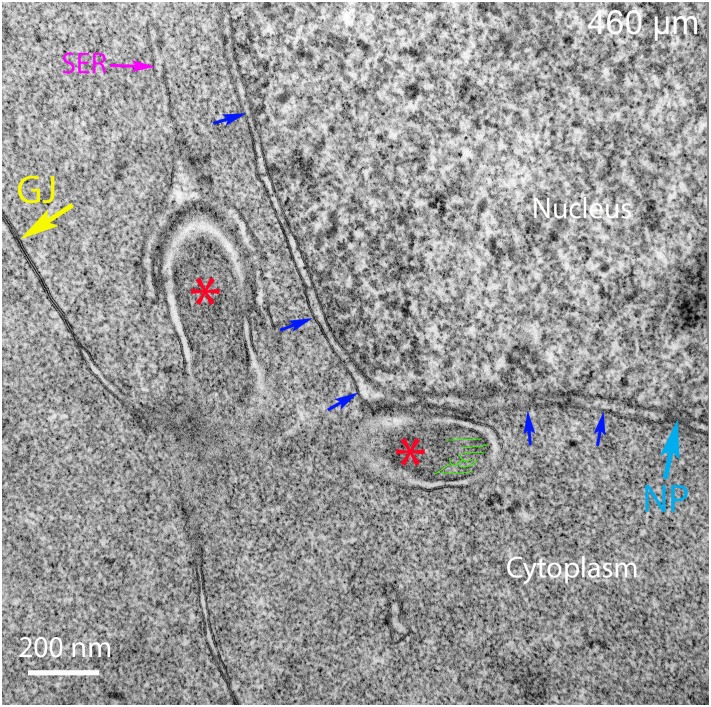
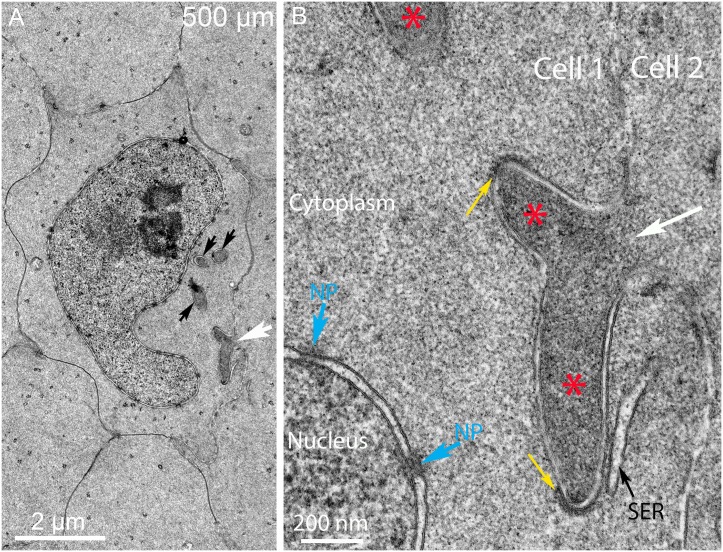
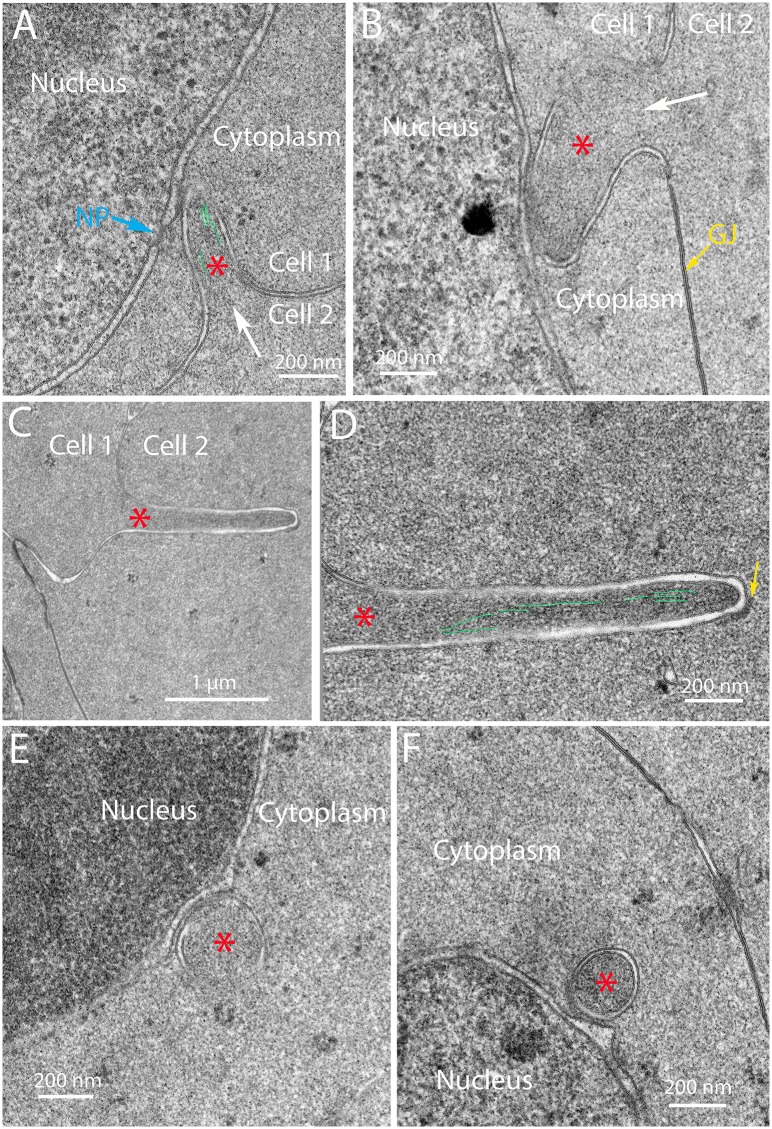
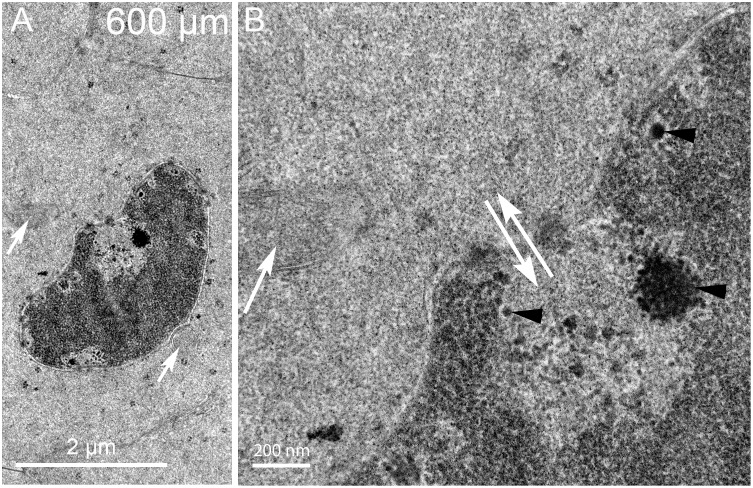
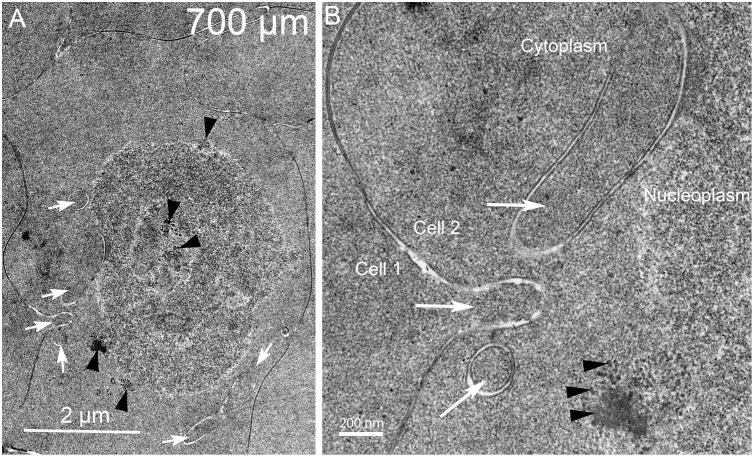
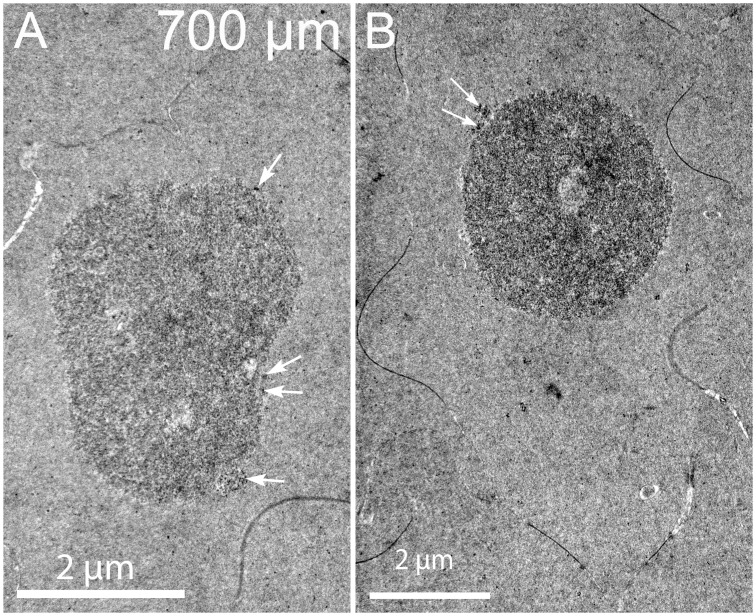
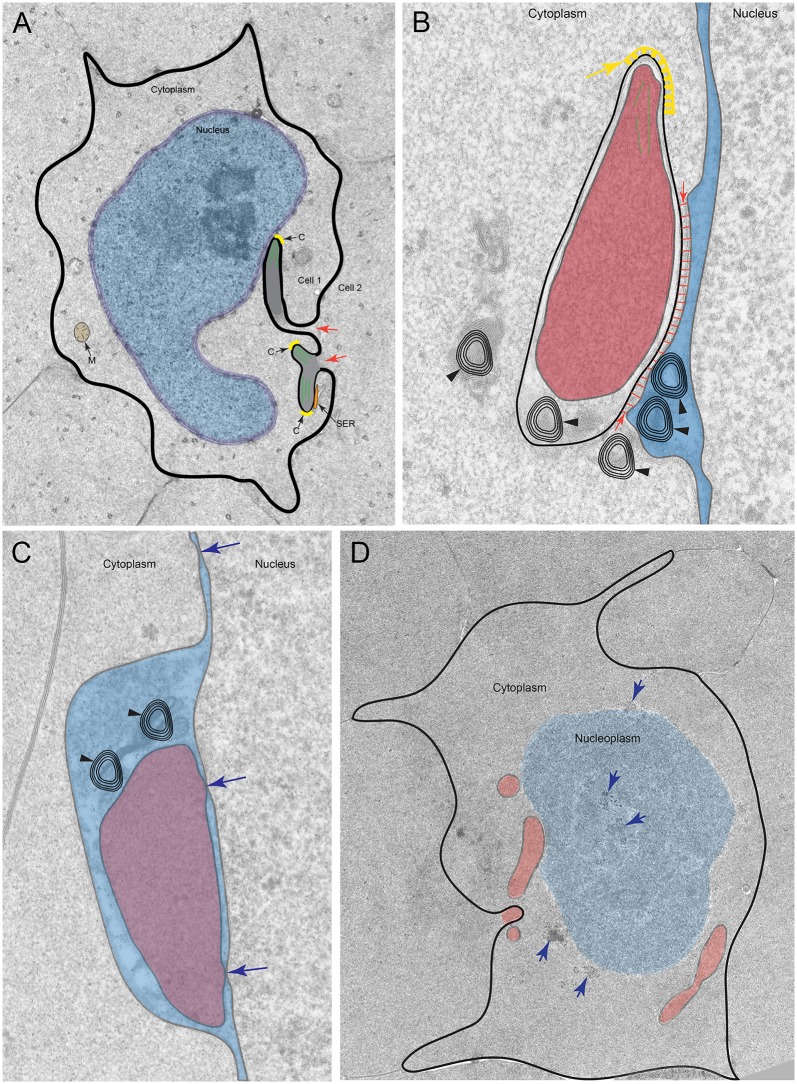
An unresolved issue in structural biology is how the encapsulated lens removes membranous organelles to carry out its role as a transparent optical element. In this ultrastructural study, we establish a mechanism for nuclear elimination in the developing chick lens during the formation of the organelle-free zone. Day 12-15 chick embryo lenses were examined by high-resolution confocal light microscopy and thin section transmission electron microscopy (TEM) following fixation in 10% formalin and 4% paraformaldehyde, and then processing for confocal or TEM as described previously. Examination of developing fiber cells revealed normal nuclei with dispersed chromatin and clear nucleoli typical of cells in active ribosome production to support protein synthesis. Early signs of nuclear degradation were observed about 300 μm from the lens capsule in Day 15 lenses where the nuclei display irregular nuclear stain and prominent indentations that sometimes contained a previously undescribed macromolecular aggregate attached to the nuclear envelope. We have termed this novel structure the nuclear excisosome. This complex by confocal is closely adherent to the nuclear envelope and by TEM appears to degrade the outer leaflet of the nuclear envelope, then the inner leaflet up to 500 μm depth. The images suggest that the nuclear excisosome separates nuclear membrane proteins from lipids, which then form multilamellar assemblies that stain intensely in confocal and in TEM have 5 nm spacing consistent with pure lipid bilayers. The denuded nucleoplasm then degrades by condensation and loss of structure in the range 600 to 700 μm depth producing pyknotic nuclear remnants. None of these stages display any classic autophagic vesicles or lysosomes associated with nuclei. Uniquely, the origin of the nuclear excisosome is from filopodial-like projections of adjacent lens fiber cells that initially contact, and then appear to fuse with the outer nuclear membrane. These filopodial-like projections appear to be initiated with a clathrin-like coat and driven by an internal actin network. In summary, a specialized cellular organelle, the nuclear excisosome, generated in part by adjacent fiber cells degrades nuclei during fiber cell differentiation and maturation.
Conflict of interest statement
Figures

Similar articles
-
Novel mitochondrial derived Nuclear Excisosome degrades nuclei during differentiation of prosimian Galago (bush baby) monkey lenses.PLoS One. 2020 Nov 12;15(11):e0241631. doi: 10.1371/journal.pone.0241631. eCollection 2020. PLoS One. 2020. PMID: 33180800 Free PMC article.
-
Chromatin degradation in differentiating fiber cells of the eye lens.J Cell Biol. 1997 Apr 7;137(1):37-49. doi: 10.1083/jcb.137.1.37. J Cell Biol. 1997. PMID: 9105035 Free PMC article.
-
Identification and expression of Hop, an atypical homeobox gene expressed late in lens fiber cell terminal differentiation.Mol Vis. 2007 Jan 26;13:114-24. Mol Vis. 2007. PMID: 17277742 Free PMC article.
-
Autophagy Requirements for Eye Lens Differentiation and Transparency.Cells. 2023 Feb 1;12(3):475. doi: 10.3390/cells12030475. Cells. 2023. PMID: 36766820 Free PMC article. Review.
-
Disassembly of the lens fiber cell nucleus to create a clear lens: The p27 descent.Exp Eye Res. 2017 Mar;156:72-78. doi: 10.1016/j.exer.2016.02.011. Epub 2016 Mar 3. Exp Eye Res. 2017. PMID: 26946072 Free PMC article. Review.
Cited by
-
Spatial Analysis of Single Fiber Cells of the Developing Ocular Lens Reveals Regulated Heterogeneity of Gene Expression.iScience. 2018 Dec 21;10:66-79. doi: 10.1016/j.isci.2018.11.024. Epub 2018 Nov 17. iScience. 2018. PMID: 30508719 Free PMC article.
-
Differential Eye Expression of Xenopus Acyltransferase Gnpat and Its Biochemical Characterization Shed Light on Lipid-Associated Ocular Pathologies.Invest Ophthalmol Vis Sci. 2023 May 1;64(5):17. doi: 10.1167/iovs.64.5.17. Invest Ophthalmol Vis Sci. 2023. PMID: 37204785 Free PMC article.
-
Uncovering mechanisms of nuclear degradation in keratinocytes: A paradigm for nuclear degradation in other tissues.Nucleus. 2018 Jan 1;9(1):56-64. doi: 10.1080/19491034.2017.1412027. Epub 2018 Jan 3. Nucleus. 2018. PMID: 29205081 Free PMC article. Review.
-
Dysregulation of Autophagy Occurs During Congenital Cataract Development in βA3ΔG91 Mice.Invest Ophthalmol Vis Sci. 2024 Apr 1;65(4):4. doi: 10.1167/iovs.65.4.4. Invest Ophthalmol Vis Sci. 2024. PMID: 38558092 Free PMC article.
-
Signaling and Gene Regulatory Networks in Mammalian Lens Development.Trends Genet. 2017 Oct;33(10):677-702. doi: 10.1016/j.tig.2017.08.001. Epub 2017 Aug 31. Trends Genet. 2017. PMID: 28867048 Free PMC article. Review.
References
-
- Kuszak JR, Brown HG. Embryology and Anatomy of the Lens In: Alberts DM, Jakobiec FA, editors. Principles and Practice of Ophthalmology. Philadelphia: W. B. Saunders; 1994. pp. 82–96.
-
- Kuszak JR, Costello MJ. Structure of the vertebrate lens In: Lovicu FJ, Robinson ML, editors. Development of the Ocular Lens. Cambridge, UK: Cambridge Univ. Press; 2004. pp. 71–118.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
