

Codon usage affects the structure and function of the Drosophila circadian clock protein PERIOD
- PMID: 27542830
- PMCID: PMC5002980
- DOI: 10.1101/gad.281030.116
Codon usage affects the structure and function of the Drosophila circadian clock protein PERIOD
Abstract
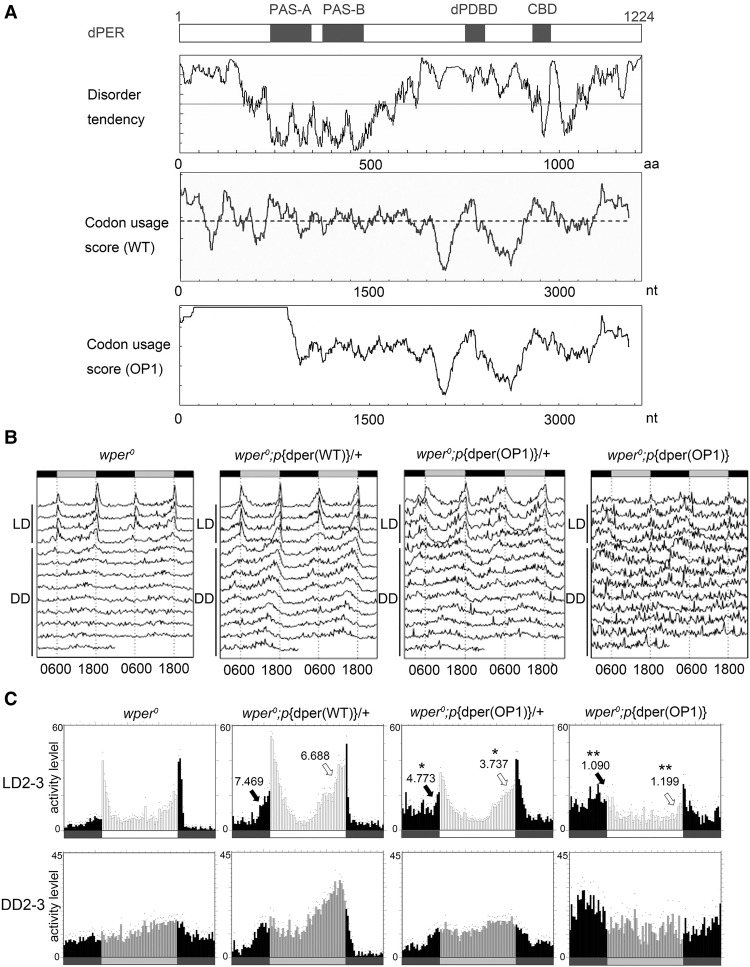
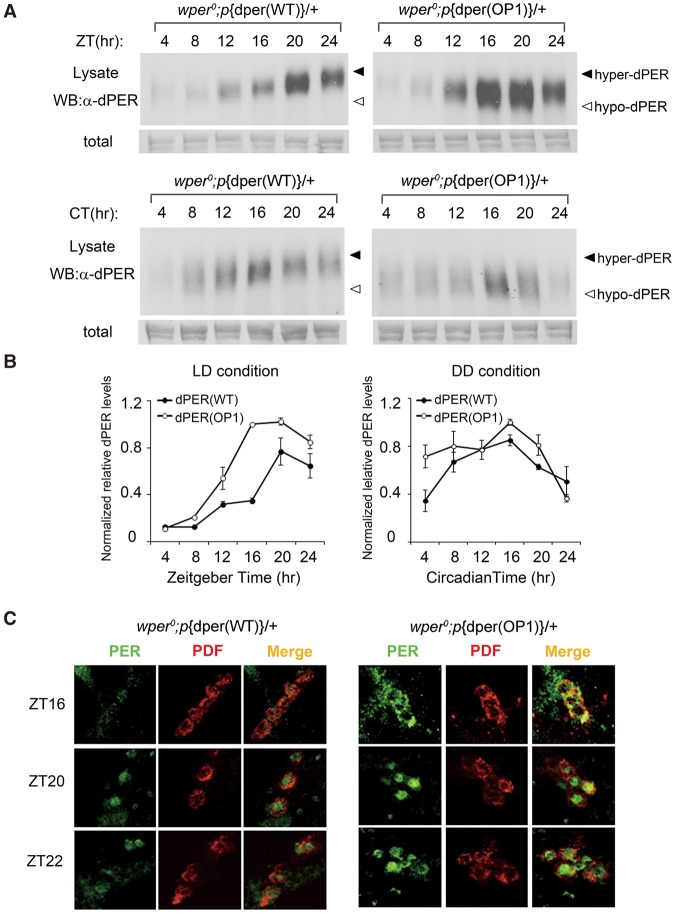
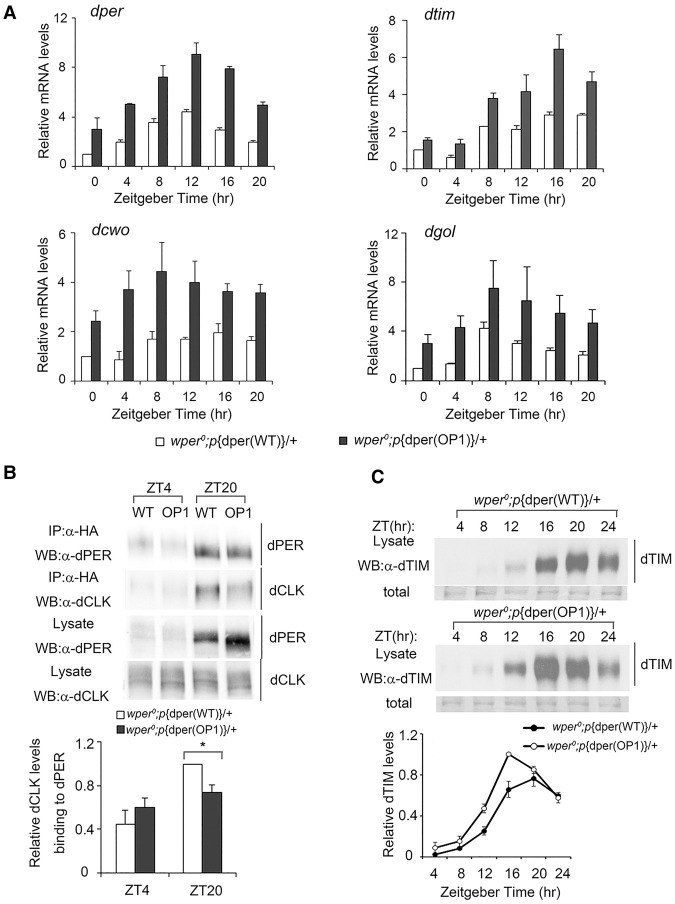
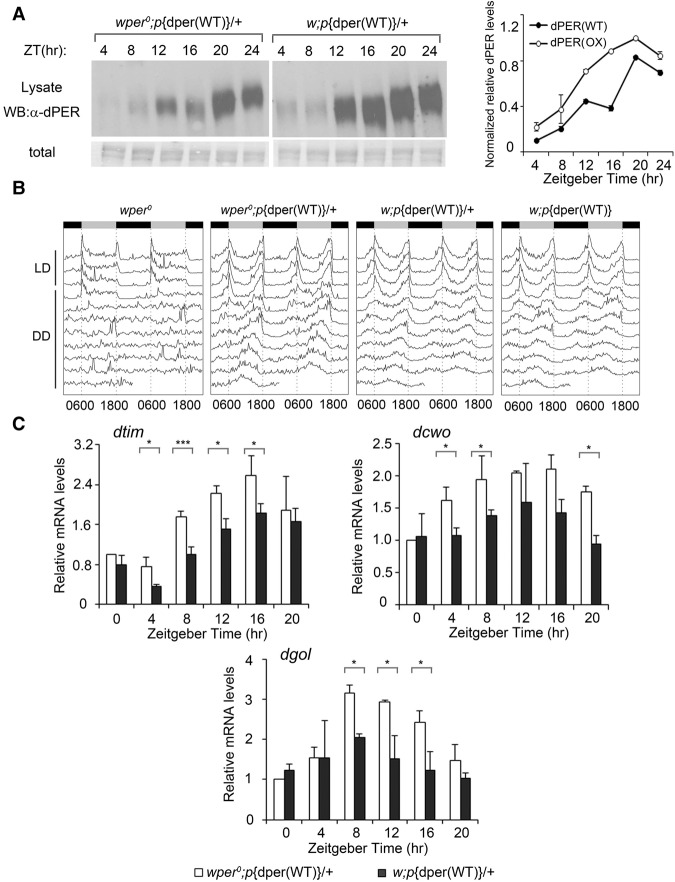
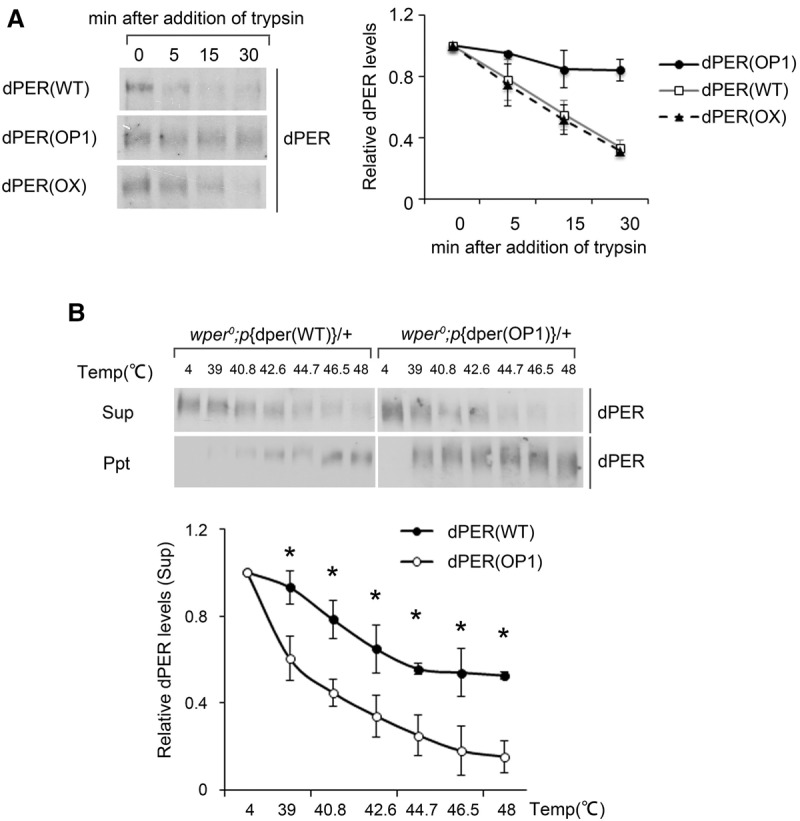
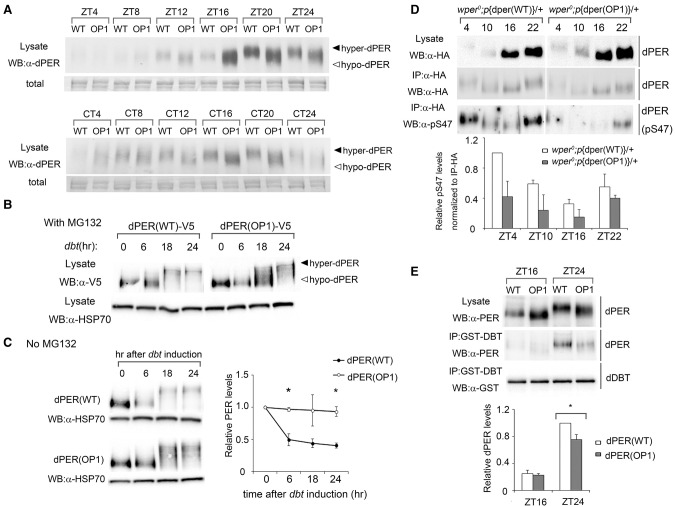
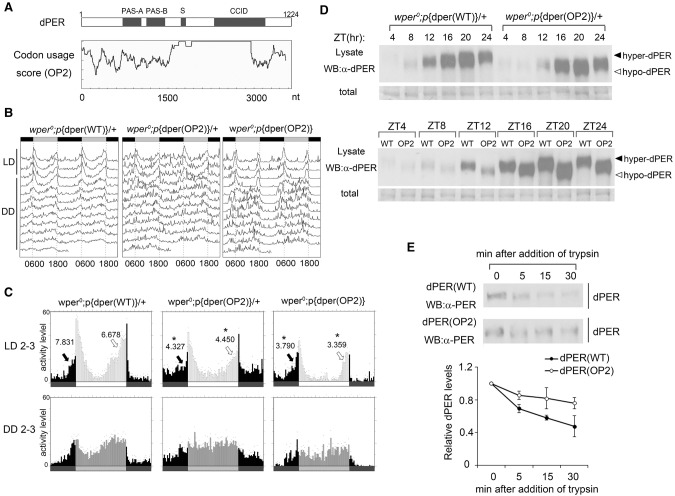
Codon usage bias is a universal feature of all genomes, but its in vivo biological functions in animal systems are not clear. To investigate the in vivo role of codon usage in animals, we took advantage of the sensitivity and robustness of the Drosophila circadian system. By codon-optimizing parts of Drosophila period (dper), a core clock gene that encodes a critical component of the circadian oscillator, we showed that dper codon usage is important for circadian clock function. Codon optimization of dper resulted in conformational changes of the dPER protein, altered dPER phosphorylation profile and stability, and impaired dPER function in the circadian negative feedback loop, which manifests into changes in molecular rhythmicity and abnormal circadian behavioral output. This study provides an in vivo example that demonstrates the role of codon usage in determining protein structure and function in an animal system. These results suggest a universal mechanism in eukaryotes that uses a codon usage "code" within genetic codons to regulate cotranslational protein folding.
Keywords: Drosophila; circadian clock; codon usage; period; protein structure.
© 2016 Fu et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Two distinct modes of PERIOD recruitment onto dCLOCK reveal a novel role for TIMELESS in circadian transcription.J Neurosci. 2010 Oct 27;30(43):14458-69. doi: 10.1523/JNEUROSCI.2366-10.2010. J Neurosci. 2010. PMID: 20980603 Free PMC article.
-
A DOUBLETIME kinase binding domain on the Drosophila PERIOD protein is essential for its hyperphosphorylation, transcriptional repression, and circadian clock function.Mol Cell Biol. 2007 Jul;27(13):5014-28. doi: 10.1128/MCB.02339-06. Epub 2007 Apr 23. Mol Cell Biol. 2007. PMID: 17452449 Free PMC article.
-
Non-optimal codon usage affects expression, structure and function of clock protein FRQ.Nature. 2013 Mar 7;495(7439):111-5. doi: 10.1038/nature11833. Epub 2013 Feb 17. Nature. 2013. PMID: 23417067 Free PMC article.
-
A hierarchical phosphorylation cascade that regulates the timing of PERIOD nuclear entry reveals novel roles for proline-directed kinases and GSK-3beta/SGG in circadian clocks.J Neurosci. 2010 Sep 22;30(38):12664-75. doi: 10.1523/JNEUROSCI.1586-10.2010. J Neurosci. 2010. PMID: 20861372 Free PMC article.
-
Circadian Control of Antibacterial Immunity: Findings from Animal Models.Front Cell Infect Microbiol. 2016 May 10;6:54. doi: 10.3389/fcimb.2016.00054. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27242972 Free PMC article. Review.
Cited by
-
Senecavirus A Enhances Its Adaptive Evolution via Synonymous Codon Bias Evolution.Viruses. 2022 May 16;14(5):1055. doi: 10.3390/v14051055. Viruses. 2022. PMID: 35632797 Free PMC article.
-
Universal protein misfolding intermediates can bypass the proteostasis network and remain soluble and less functional.Nat Commun. 2022 Jun 2;13(1):3081. doi: 10.1038/s41467-022-30548-5. Nat Commun. 2022. PMID: 35654797 Free PMC article.
-
O-GlcNAcylation of PERIOD regulates its interaction with CLOCK and timing of circadian transcriptional repression.PLoS Genet. 2019 Jan 31;15(1):e1007953. doi: 10.1371/journal.pgen.1007953. eCollection 2019 Jan. PLoS Genet. 2019. PMID: 30703153 Free PMC article.
-
The Codon Usage Code for Cotranslational Folding of Viral Capsids.Genome Biol Evol. 2021 Sep 1;13(9):evab089. doi: 10.1093/gbe/evab089. Genome Biol Evol. 2021. PMID: 33914886 Free PMC article.
-
Quantifying shifts in natural selection on codon usage between protein regions: a population genetics approach.BMC Genomics. 2022 May 30;23(1):408. doi: 10.1186/s12864-022-08635-0. BMC Genomics. 2022. PMID: 35637464 Free PMC article.
References
-
- Bae K, Edery I. 2006. Regulating a circadian clock's period, phase and amplitude by phosphorylation: insights from Drosophila. J Biochem 140: 609–617. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases