

An interplay between the serotonin transporter (SERT) and 5-HT receptors controls stimulus-secretion coupling in sympathoadrenal chromaffin cells
- PMID: 27544824
- PMCID: PMC5028315
- DOI: 10.1016/j.neuropharm.2016.08.015
An interplay between the serotonin transporter (SERT) and 5-HT receptors controls stimulus-secretion coupling in sympathoadrenal chromaffin cells
Abstract
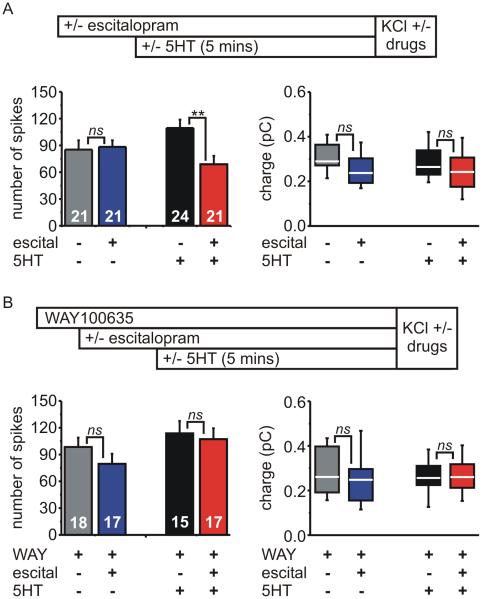
Adrenal chromaffin cells (ACCs), the neuroendocrine arm of the sympathetic nervous system, secrete catecholamines to mediate the physiological response to stress. Although ACCs do not synthesize 5-HT, they express the serotonin transporter (SERT). Genetic variations in SERT are linked to several CNS disorders but the role(s) of SERT/5-HT in ACCs has remained unclear. Adrenal glands from wild-type mice contained 5-HT at ≈ 750 fold lower abundance than adrenaline, and in SERT(-/-) mice this was reduced by ≈80% with no change in catecholamines. Carbon fibre amperometry showed that SERT modulated the ability of 5-HT1A receptors to inhibit exocytosis. 5-HT reduced the number of amperometric spikes (vesicular fusion events) evoked by KCl in SERT(-/-) cells and wild-type cells treated with escitalopram, a SERT antagonist. The 5-HT1A receptor antagonist WAY100635 blocked the inhibition by 5-HT which was mimicked by the 5-HT1A agonist 8-OH-DPAT but not the 5-HT1B agonist CP93129. There was no effect on voltage-gated Ca(2+) channels, K(+) channels, or intracellular [Ca(2+)] handling, showing the 5-HT receptors recruit an atypical inhibitory mechanism. Spike charge and kinetics were not altered by 5-HT receptors but were reduced in SERT(-/-) cells compared to wild-type cells. Our data reveal a novel role for SERT and suggest that adrenal chromaffin cells might be a previously unrecognized hub for serotonergic control of the sympathetic stress response.
Keywords: 5-HT receptor; 8-OH-DPAT (PubChem CID: 6917794); Adrenal chromaffin cell; Amperometry; CP93129 (PubChem CID: 124007); Calcium channel; Calcium imaging; Catecholamine; Escitalopram (PubChem CID: 146571); Exocytosis; GPCR; Gallein (PubChem CID: 73685); Serotonin hydrochloride (PubChem CID: 160436); Serotonin transporter; WAY100635 (PubChem CID: 11957721).
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures







Similar articles
-
Sigma-1 receptor ligands inhibit catecholamine secretion from adrenal chromaffin cells due to block of nicotinic acetylcholine receptors.J Neurochem. 2017 Oct;143(2):171-182. doi: 10.1111/jnc.14149. Epub 2017 Sep 19. J Neurochem. 2017. PMID: 28815595 Free PMC article.
-
Serotonin and Serotonin Transporters in the Adrenal Medulla: A Potential Hub for Modulation of the Sympathetic Stress Response.ACS Chem Neurosci. 2017 May 17;8(5):943-954. doi: 10.1021/acschemneuro.7b00026. Epub 2017 Apr 13. ACS Chem Neurosci. 2017. PMID: 28406285 Free PMC article. Review.
-
Adrenal serotonin derives from accumulation by the antidepressant-sensitive serotonin transporter.Pharmacol Res. 2019 Feb;140:56-66. doi: 10.1016/j.phrs.2018.06.008. Epub 2018 Jun 9. Pharmacol Res. 2019. PMID: 29894763 Free PMC article.
-
Serotonin and the serotonin transporter in the adrenal gland.Vitam Horm. 2024;124:39-78. doi: 10.1016/bs.vh.2023.06.002. Epub 2023 Jul 18. Vitam Horm. 2024. PMID: 38408804 Free PMC article. Review.
-
Enhanced Ca(2+)-induced Ca(2+) release from intracellular stores contributes to catecholamine hypersecretion in adrenal chromaffin cells from spontaneously hypertensive rats.Pflugers Arch. 2015 Nov;467(11):2307-23. doi: 10.1007/s00424-015-1702-8. Epub 2015 Mar 21. Pflugers Arch. 2015. PMID: 25791627
Cited by
-
p38α MAPK signaling drives pharmacologically reversible brain and gastrointestinal phenotypes in the SERT Ala56 mouse.Proc Natl Acad Sci U S A. 2018 Oct 23;115(43):E10245-E10254. doi: 10.1073/pnas.1809137115. Epub 2018 Oct 8. Proc Natl Acad Sci U S A. 2018. PMID: 30297392 Free PMC article.
-
Jedi-1 deficiency increases sensory neuron excitability through a non-cell autonomous mechanism.Sci Rep. 2020 Jan 28;10(1):1300. doi: 10.1038/s41598-020-57971-2. Sci Rep. 2020. PMID: 31992767 Free PMC article.
-
Restraint Stress Disrupted Intestinal Homeostasis via 5-HT/HTR7/Wnt/β-Catenin/NF-kB Signaling.Int J Mol Sci. 2025 Apr 24;26(9):4021. doi: 10.3390/ijms26094021. Int J Mol Sci. 2025. PMID: 40362261 Free PMC article.
-
Serotonergic Modulation of Neurovascular Transmission: A Focus on Prejunctional 5-HT Receptors/Mechanisms.Biomedicines. 2023 Jun 29;11(7):1864. doi: 10.3390/biomedicines11071864. Biomedicines. 2023. PMID: 37509503 Free PMC article. Review.
-
Sigma-1 receptor ligands inhibit catecholamine secretion from adrenal chromaffin cells due to block of nicotinic acetylcholine receptors.J Neurochem. 2017 Oct;143(2):171-182. doi: 10.1111/jnc.14149. Epub 2017 Sep 19. J Neurochem. 2017. PMID: 28815595 Free PMC article.
References
-
- Bayliss DA, Li YW, Talley EM. Effects of serotonin on caudal raphe neurons: activation of an inwardly rectifying potassium conductance. J Neurophysiol. 1997a;77:1349–1361. - PubMed
-
- Bayliss DA, Li YW, Talley EM. Effects of serotonin on caudal raphe neurons: inhibition of N- and P/Q-type calcium channels and the afterhyperpolarization. J Neurophysiol. 1997b;77:1362–1374. - PubMed
-
- Bengel D, Murphy DL, Andrews AM, Wichems CH, Feltner D, Heils A, Mossner R, Westphal H, Lesch KP. Altered brain serotonin homeostasis and locomotor insensitivity to 3, 4-methylenedioxymethamphetamine ("Ecstasy") in serotonin transporter-deficient mice. Mol Pharmacol. 1998;53:649–655. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
