

ALS Mutations Disrupt Phase Separation Mediated by α-Helical Structure in the TDP-43 Low-Complexity C-Terminal Domain
- PMID: 27545621
- PMCID: PMC5014597
- DOI: 10.1016/j.str.2016.07.007
ALS Mutations Disrupt Phase Separation Mediated by α-Helical Structure in the TDP-43 Low-Complexity C-Terminal Domain
Abstract
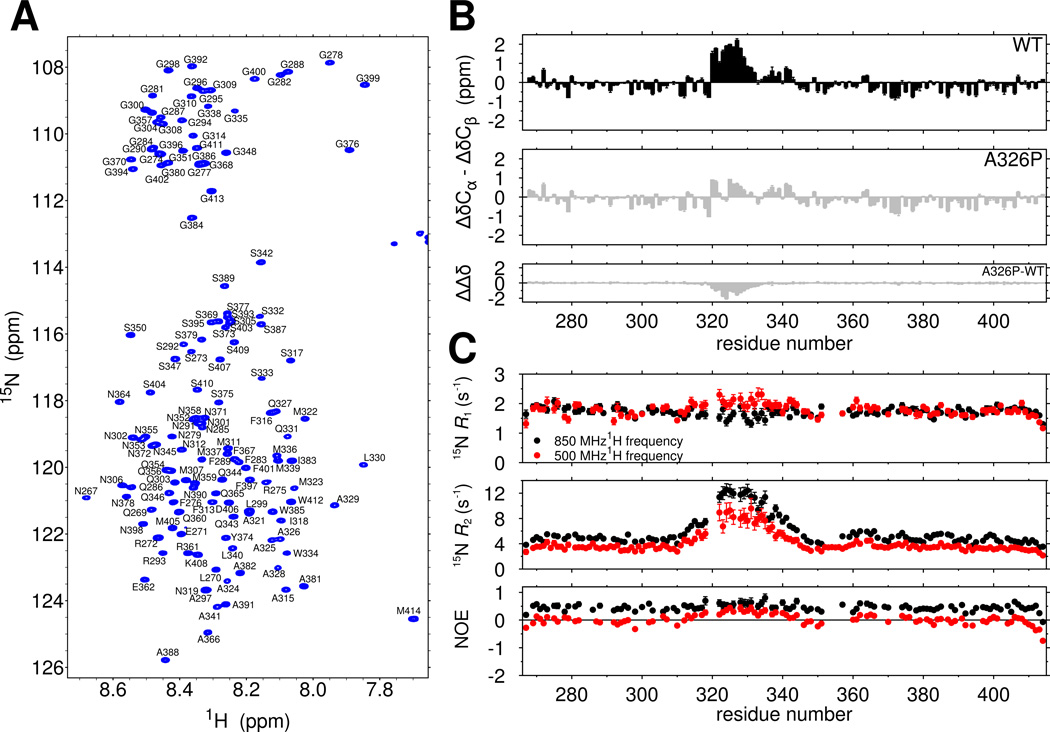
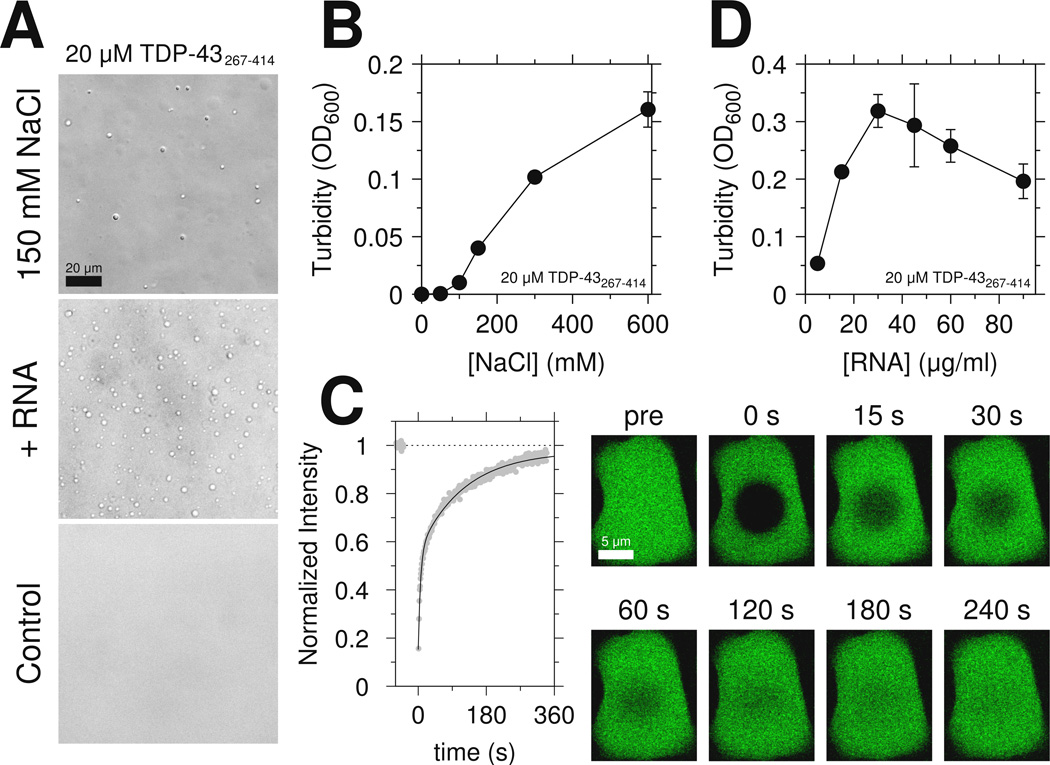
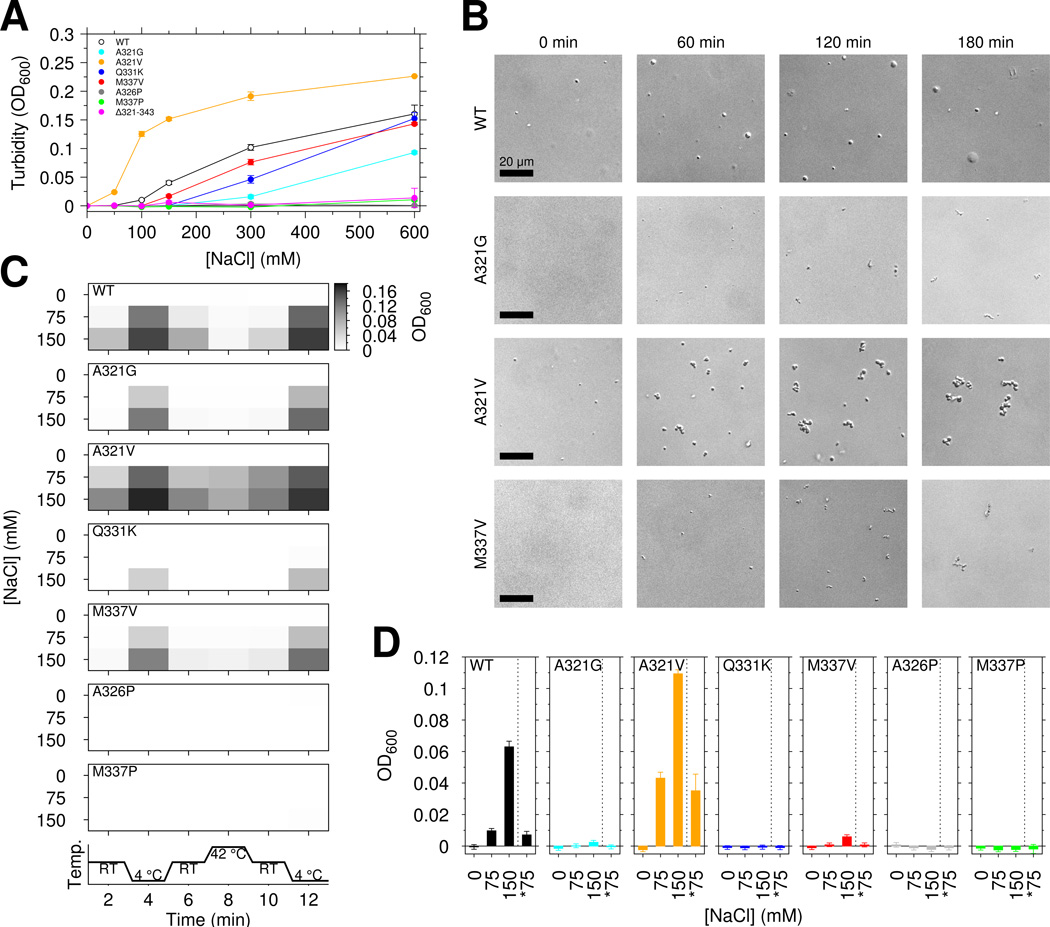
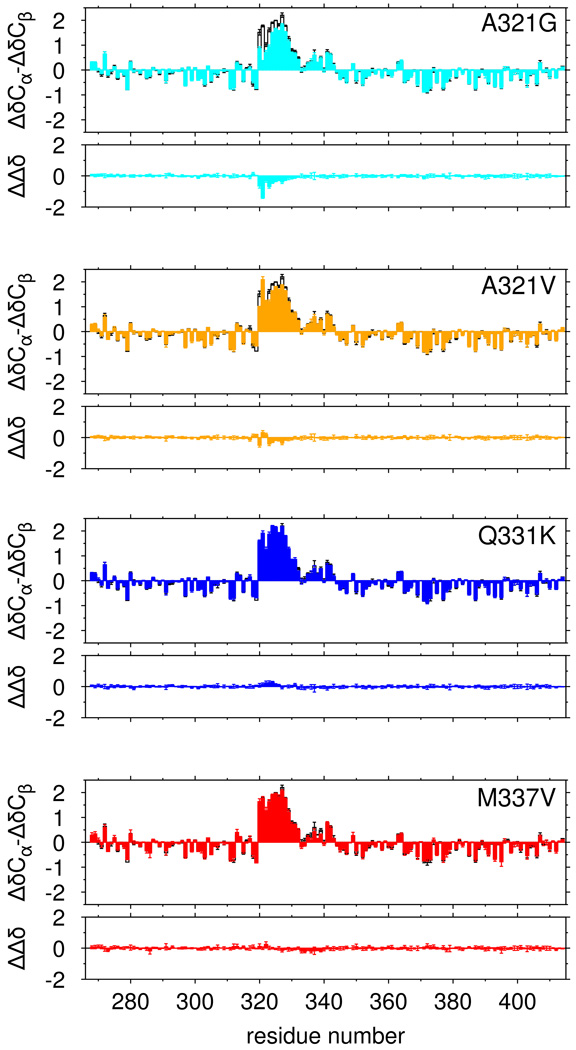
RNA-binding protein TDP-43 mediates essential RNA processing but forms cytoplasmic neuronal inclusions via its C-terminal domain (CTD) in amyotrophic lateral sclerosis (ALS). It remains unclear if aggregated TDP-43 is neurotoxic and if ∼50 ALS-associated missense mutations in TDP-43 CTD promote aggregation, or if loss of normal function plays a role in disease. Recent work points to the ability of related proteins to assemble into functional phase-separated ribonucleoprotein granules via their structurally disordered prion-like domains. Here, we provide atomic details on the structure and assembly of the low-complexity CTD of TDP-43 into liquid-liquid phase-separated in vitro granules and demonstrate that ALS-associated variants disrupt interactions within granules. Using nuclear magnetic resonance spectroscopy, simulation, and microscopy, we find that a subregion cooperatively but transiently folds into a helix that mediates TDP-43 phase separation. ALS-associated mutations disrupt phase separation by inhibiting interaction and helical stabilization. Therefore, ALS-associated mutations can disrupt TDP-43 interactions, affecting function beyond encouraging aggregation.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures

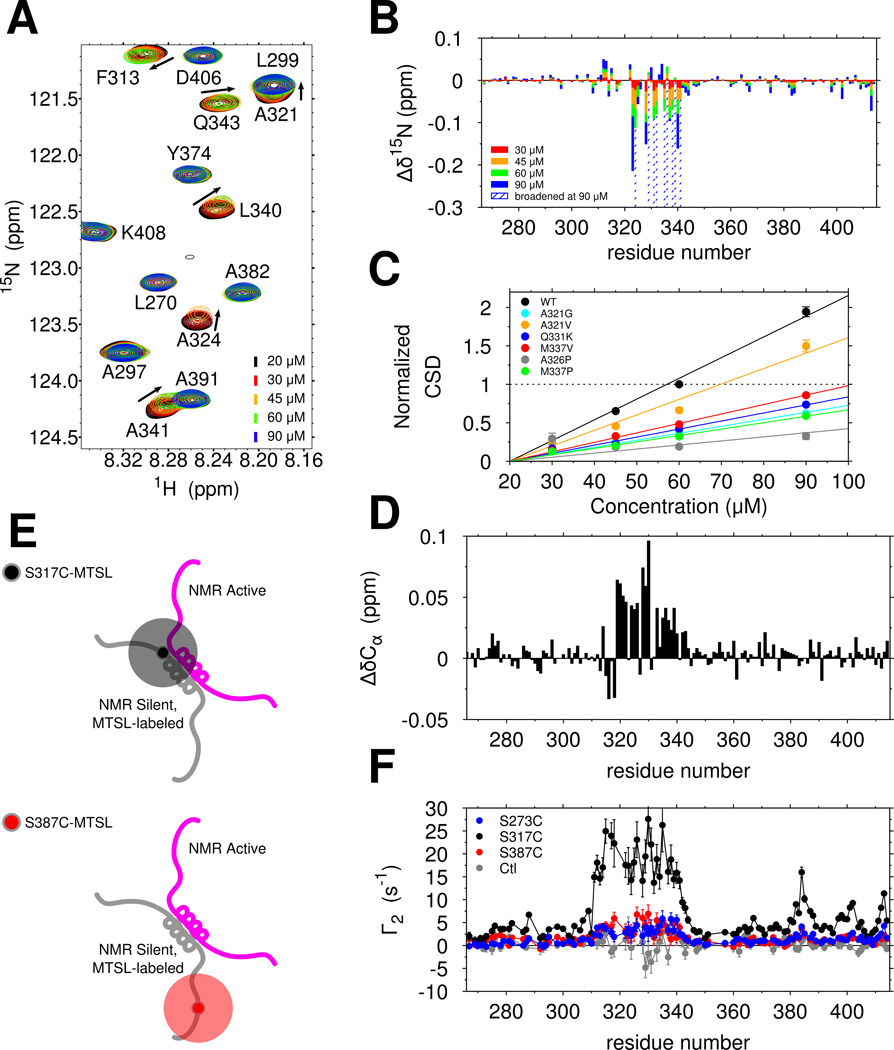
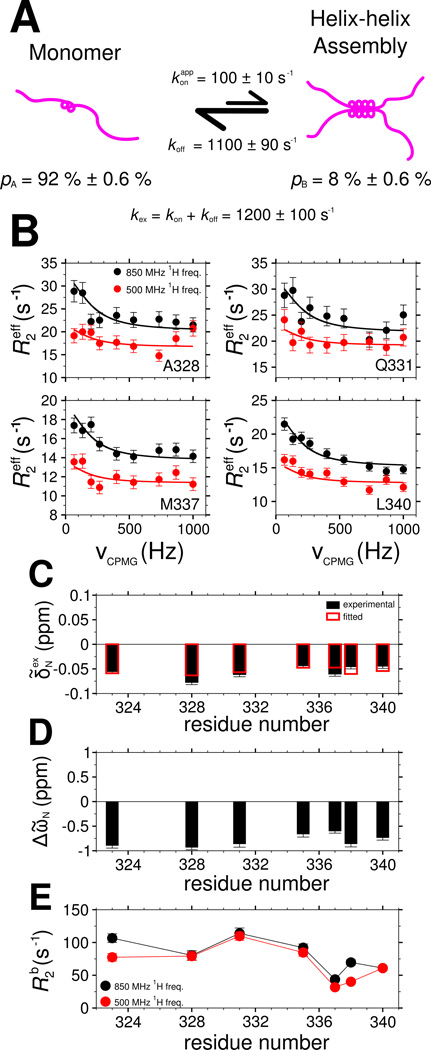
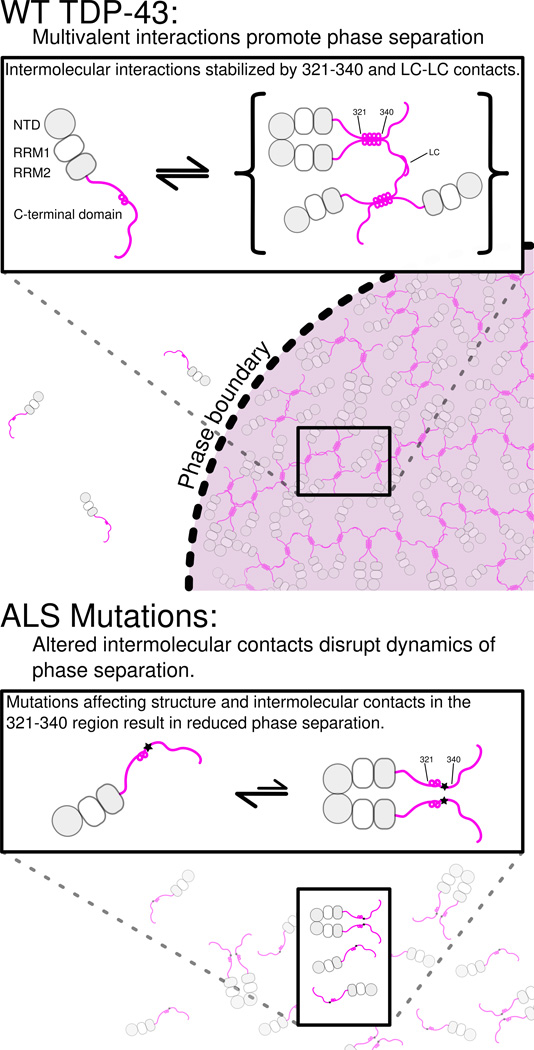
Comment in
-
A New Phase in ALS Research.Structure. 2016 Sep 6;24(9):1435-6. doi: 10.1016/j.str.2016.08.003. Structure. 2016. PMID: 27602988
References
-
- Arnold ES, Ling SC, Huelga SC, Lagier-Tourenne C, Polymenidou M, Ditsworth D, Kordasiewicz HB, McAlonis-Downes M, Platoshyn O, Parone PA, et al. ALS-linked TDP-43 mutations produce aberrant RNA splicing and adult-onset motor neuron disease without aggregation or loss of nuclear TDP-43. Proc Natl Acad Sci U S A. 2013 - PMC - PubMed
-
- Ayala YM, Pantano S, D'Ambrogio A, Buratti E, Brindisi A, Marchetti C, Romano M, Baralle FE. Human, Drosophila, and C.elegans TDP43: nucleic acid binding properties and splicing regulatory function. J Mol Biol. 2005;348:575–588. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
