

Developmental environment mediates male seminal protein investment in Drosophila melanogaster
- PMID: 27546947
- PMCID: PMC4974917
- DOI: 10.1111/1365-2435.12515
Developmental environment mediates male seminal protein investment in Drosophila melanogaster
Abstract
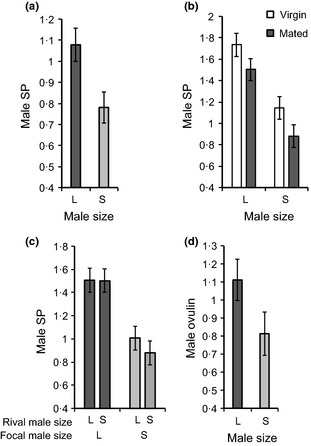
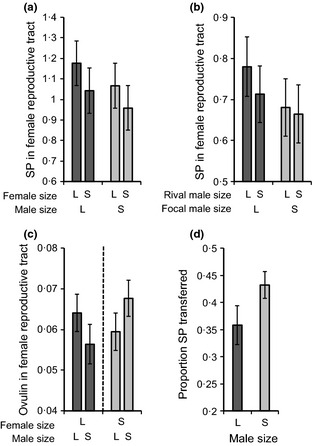
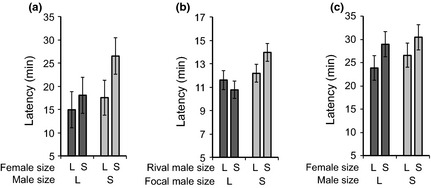
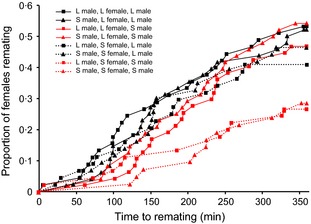
Males of many species fine-tune their ejaculates in response to sperm competition risk. Resource availability and the number of competitors during development can also strongly influence sperm production. However, despite the key role of seminal proteins in mediating reproductive processes, it is unclear whether seminal protein investment is dependent on the developmental environment.We manipulated the developmental environment of Drosophila melanogaster by rearing flies at low and high density. As expected, this resulted in large and small (i.e. high and low condition) adult phenotypes, respectively.As predicted, large males produced more of two key seminal proteins, sex peptide (SP) and ovulin, and were more successful at obtaining matings with both virgin and previously mated females. However, there was only a weak and non-significant trend for large males to transfer more absolute quantities of SP at mating, and thus, small males ejaculated proportionally more of their stored accessory gland SP resources.Males transferred more receptivity-inhibiting SP to large females. Despite this, large females remated more quickly than small females and thus responded to their developmental environment over and above the quantity of SP they received.The results are consistent with two non-mutually exclusive hypotheses. First, flies might respond to condition-dependent reproductive opportunities, with (i) small males investing heavily in ejaculates when mating opportunities arise and large males strategically partitioning SP resources and (ii) small females remating at reduced rates because they have higher mating costs or need to replenish sperm less often.Second, flies may be primed by their larval environment to deal with similar adult population densities, with (i) males perceiving high density as signalling increased competition, leading small males to invest proportionally more SP resources at mating and (ii) females perceiving high density as signalling abundant potential mates, leading to a higher sexual receptivity threshold.Thus, by influencing the mating frequencies of both sexes, as well as the quantity of seminal proteins produced by males and received by females, the developmental environment is likely to have far-reaching and sex-specific consequences for sexual selection and sexual conflict.
Keywords: condition; density; development; ovulin; sex peptide; sexual selection.
Figures
Similar articles
-
The developmental environment modulates mating-induced aggression and fighting success in adult female Drosophila.Funct Ecol. 2018 Nov;32(11):2542-2552. doi: 10.1111/1365-2435.13214. Epub 2018 Sep 28. Funct Ecol. 2018. PMID: 31007331 Free PMC article.
-
Protein-specific manipulation of ejaculate composition in response to female mating status in Drosophila melanogaster.Proc Natl Acad Sci U S A. 2011 Jun 14;108(24):9922-6. doi: 10.1073/pnas.1100905108. Epub 2011 May 31. Proc Natl Acad Sci U S A. 2011. PMID: 21628597 Free PMC article.
-
The benefits of male ejaculate sex peptide transfer in Drosophila melanogaster.J Evol Biol. 2009 Feb;22(2):275-86. doi: 10.1111/j.1420-9101.2008.01638.x. Epub 2008 Nov 15. J Evol Biol. 2009. PMID: 19032499
-
Female remating, sperm competition and sexual selection in Drosophila.Genet Mol Res. 2002 Sep 30;1(3):178-215. Genet Mol Res. 2002. PMID: 14963827 Review.
-
The gifts that keep on giving: physiological functions and evolutionary dynamics of male seminal proteins in Drosophila.Heredity (Edinb). 2002 Feb;88(2):85-93. doi: 10.1038/sj.hdy.6800017. Heredity (Edinb). 2002. PMID: 11932766 Review.
Cited by
-
A predominant role of genotypic variation in both expression of sperm competition genes and paternity success in Drosophila melanogaster.Proc Biol Sci. 2023 Sep 27;290(2007):20231715. doi: 10.1098/rspb.2023.1715. Epub 2023 Sep 20. Proc Biol Sci. 2023. PMID: 37727083 Free PMC article.
-
Crowded developmental environment promotes adult sex-specific nutrient consumption in a polyphagous fly.Front Zool. 2019 Feb 18;16:4. doi: 10.1186/s12983-019-0302-4. eCollection 2019. Front Zool. 2019. PMID: 30820236 Free PMC article.
-
Epigenetic paternal effects as costly, condition-dependent traits.Heredity (Edinb). 2018 Sep;121(3):248-256. doi: 10.1038/s41437-018-0096-8. Epub 2018 Jun 14. Heredity (Edinb). 2018. PMID: 29904169 Free PMC article. Review.
-
The developmental environment modulates mating-induced aggression and fighting success in adult female Drosophila.Funct Ecol. 2018 Nov;32(11):2542-2552. doi: 10.1111/1365-2435.13214. Epub 2018 Sep 28. Funct Ecol. 2018. PMID: 31007331 Free PMC article.
-
The evolution of sex peptide: sexual conflict, cooperation, and coevolution.Biol Rev Camb Philos Soc. 2022 Aug;97(4):1426-1448. doi: 10.1111/brv.12849. Epub 2022 Mar 6. Biol Rev Camb Philos Soc. 2022. PMID: 35249265 Free PMC article.
References
-
- Alonzo, S.H. & Pizzari, T. (2010) Male fecundity stimulation: conflict and cooperation within and between the sexes: model analyses and coevolutionary dynamics. The American Naturalist, 175, 174–185. - PubMed
-
- Amitin, E.G. & Pitnick, S. (2007) Influence of developmental environment on male‐ and female‐mediated sperm precedence in Drosophila melanogaster . Journal of Evolutionary Biology, 20, 381–391. - PubMed
-
- Bangham, J. , Partridge, L. & Chapman, T. (2002) Effects of body size, accessory gland and testis size on pre‐ and postcopulatory success in Drosophila melanogaster . Animal Behaviour, 64, 915–921.
-
- Birkhead, T. , Hosken, D. & Pitnick, S. (2008) Sperm Biology: An Evolutionary Perspective. Academic Press, Waltham, Mass.
-
- Bolker, B.M. , Brooks, M.E. , Clark, C.J. , Geange, S.W. , Poulsen, J.R. , Stevens, M.H.H. et al (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology & Evolution, 24, 127–135. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials