

Aggregation and Prion-Like Properties of Misfolded Tumor Suppressors: Is Cancer a Prion Disease?
- PMID: 27549118
- PMCID: PMC5046694
- DOI: 10.1101/cshperspect.a023614
Aggregation and Prion-Like Properties of Misfolded Tumor Suppressors: Is Cancer a Prion Disease?
Abstract
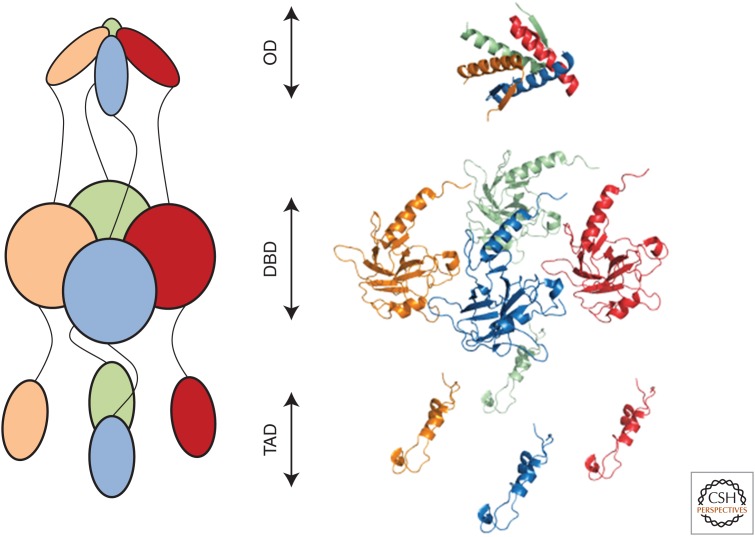
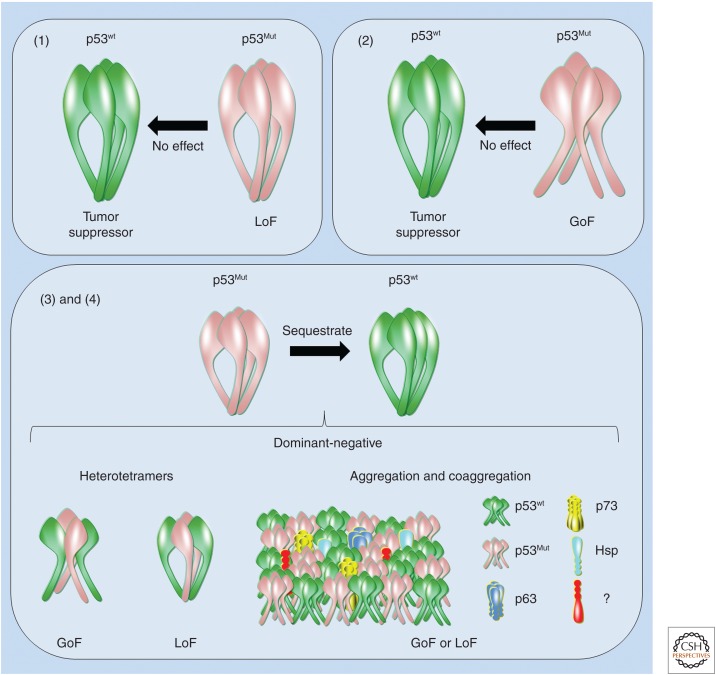
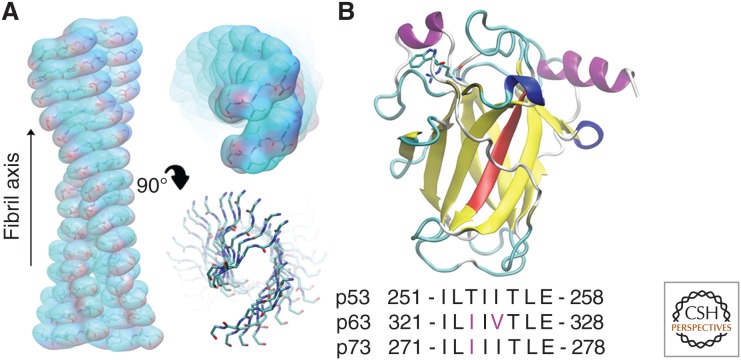
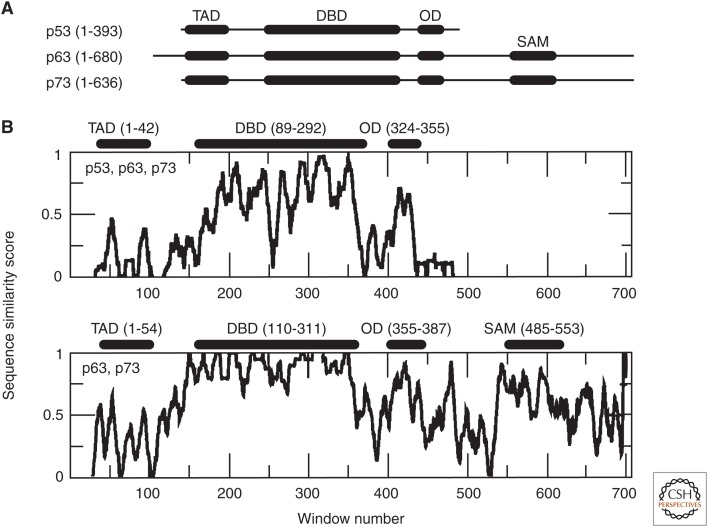
Prion diseases are disorders that share several characteristics that are typical of many neurodegenerative diseases. Recently, several studies have extended the prion concept to pathological aggregation in malignant tumors involving misfolded p53, a tumor-suppressor protein. The aggregation of p53 and its coaggregation with p53 family members, p63 and p73, have been shown. Certain p53 mutants exert a dominant-negative regulatory effect on wild-type (WT) p53. The basis for this dominant-negative effect is that amyloid-like mutant p53 converts WT p53 into an aggregated species, leading to a gain-of-function (GoF) phenotype and the loss of its tumor-suppressor function. Recently, it was shown that p53 aggregates can be internalized by cells and can coaggregate with endogenous p53, corroborating the prion-like properties of p53 aggregates. The prion-like behavior of oncogenic p53 mutants provides an explanation for its dominant-negative and GoF properties, including the high metastatic potential of cancer cells carrying p53 mutations. The inhibition of p53 aggregation appears to represent a promising target for therapeutic intervention in patients with malignant tumors.
Copyright © 2016 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
Similar articles
-
Phase Separation and Prion-Like Aggregation of p53 Family Tumor Suppressors: From Protein Evolution to Cancer Treatment.J Neurochem. 2025 Apr;169(4):e70055. doi: 10.1111/jnc.70055. J Neurochem. 2025. PMID: 40178008 Review.
-
The aggregation of mutant p53 produces prion-like properties in cancer.Prion. 2014 Jan-Feb;8(1):75-84. doi: 10.4161/pri.27776. Prion. 2014. PMID: 24509441 Free PMC article.
-
Prion-like aggregation of mutant p53 in cancer.Trends Biochem Sci. 2014 Jun;39(6):260-7. doi: 10.1016/j.tibs.2014.04.001. Epub 2014 Apr 26. Trends Biochem Sci. 2014. PMID: 24775734 Review.
-
Expanding the prion concept to cancer biology: dominant-negative effect of aggregates of mutant p53 tumour suppressor.Biosci Rep. 2013 Jul 25;33(4):e00054. doi: 10.1042/BSR20130065. Biosci Rep. 2013. PMID: 24003888 Free PMC article. Review.
-
Targeting the Prion-like Aggregation of Mutant p53 to Combat Cancer.Acc Chem Res. 2018 Jan 16;51(1):181-190. doi: 10.1021/acs.accounts.7b00473. Epub 2017 Dec 20. Acc Chem Res. 2018. PMID: 29260852 Review.
Cited by
-
Amyloid aggregates induced by the p53-R280T mutation lead to loss of p53 function in nasopharyngeal carcinoma.Cell Death Dis. 2024 Jan 11;15(1):35. doi: 10.1038/s41419-024-06429-8. Cell Death Dis. 2024. PMID: 38212344 Free PMC article.
-
The interplay between mutant p53 and the mevalonate pathway.Cell Death Differ. 2018 Mar;25(3):460-470. doi: 10.1038/s41418-017-0026-y. Epub 2017 Dec 13. Cell Death Differ. 2018. PMID: 29238070 Free PMC article. Review.
-
Nucleo-cytoplasmic environment modulates spatiotemporal p53 phase separation.Sci Adv. 2024 Dec 13;10(50):eads0427. doi: 10.1126/sciadv.ads0427. Epub 2024 Dec 11. Sci Adv. 2024. PMID: 39661689 Free PMC article.
-
Aggregation-primed molten globule conformers of the p53 core domain provide potential tools for studying p53C aggregation in cancer.J Biol Chem. 2018 Jul 20;293(29):11374-11387. doi: 10.1074/jbc.RA118.003285. Epub 2018 May 31. J Biol Chem. 2018. PMID: 29853637 Free PMC article.
-
Aminoquinolones and Their Benzoquinone Dimer Hybrids as Modulators of Prion Protein Conversion.Molecules. 2022 Nov 16;27(22):7935. doi: 10.3390/molecules27227935. Molecules. 2022. PMID: 36432036 Free PMC article.
References
-
- Ahn J, Prives C. 2001. The C-terminus of p53: The more you learn the less you know. Nat Struct Biol 8: 730–732. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous