

Glucose enhances indolic glucosinolate biosynthesis without reducing primary sulfur assimilation
- PMID: 27549907
- PMCID: PMC4994012
- DOI: 10.1038/srep31854
Glucose enhances indolic glucosinolate biosynthesis without reducing primary sulfur assimilation
Abstract
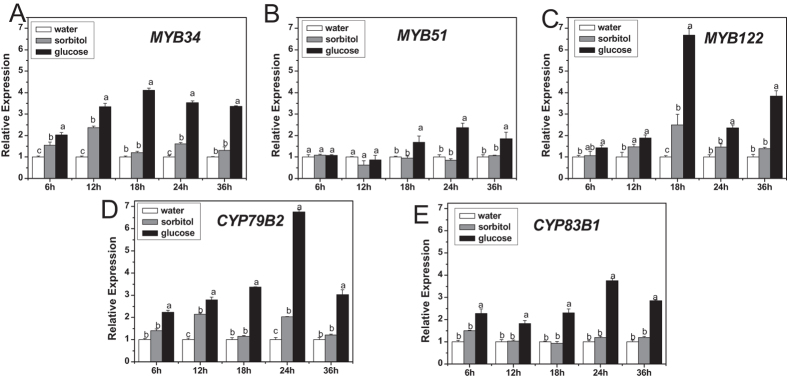
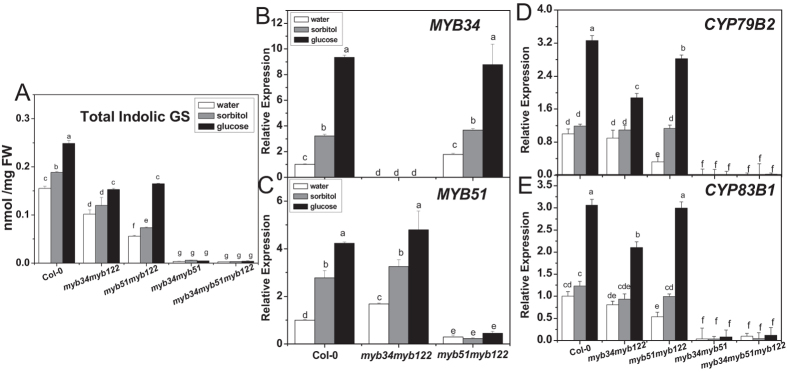
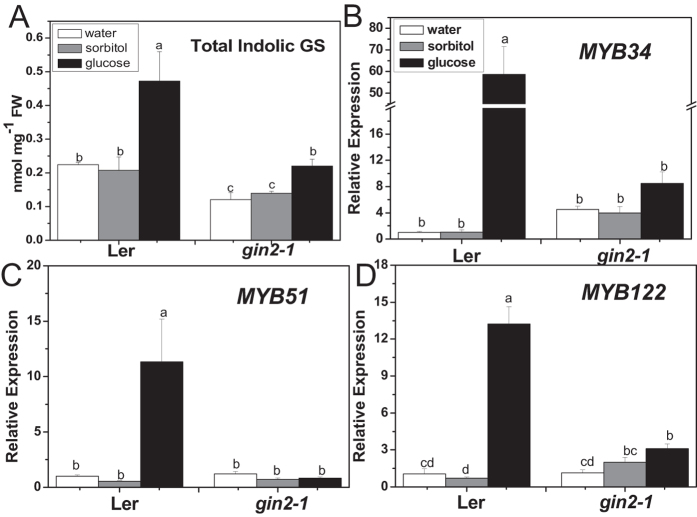
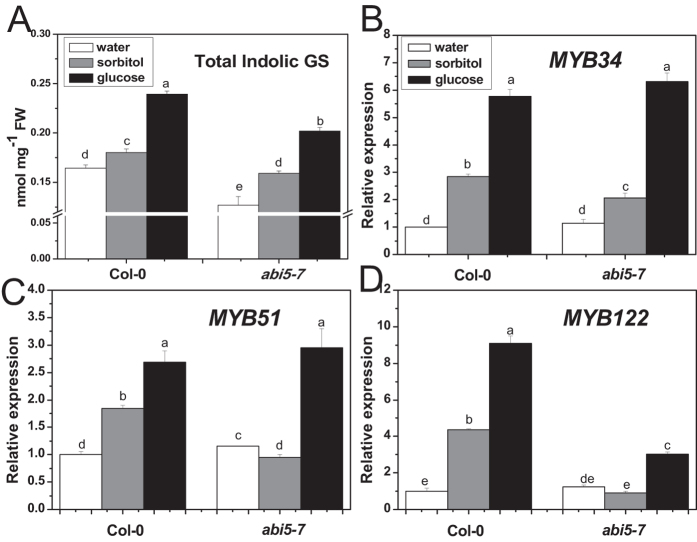
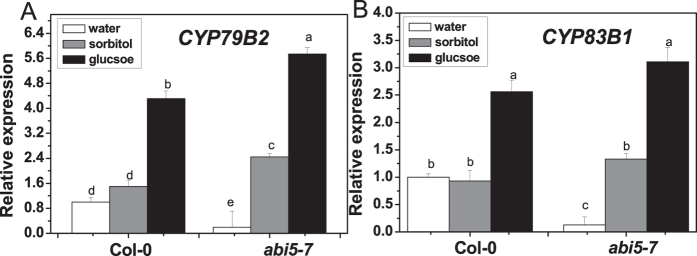
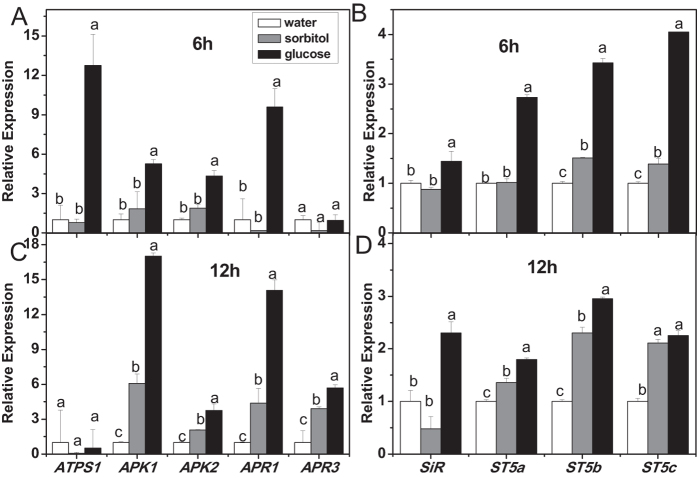
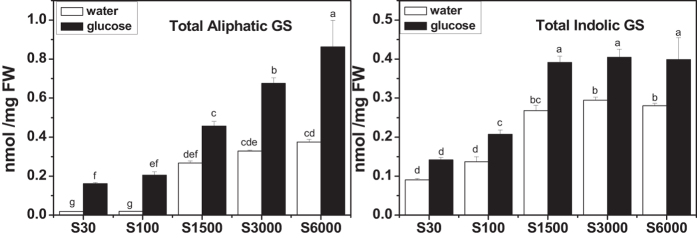
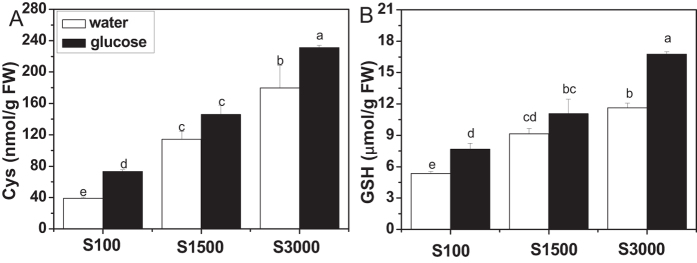
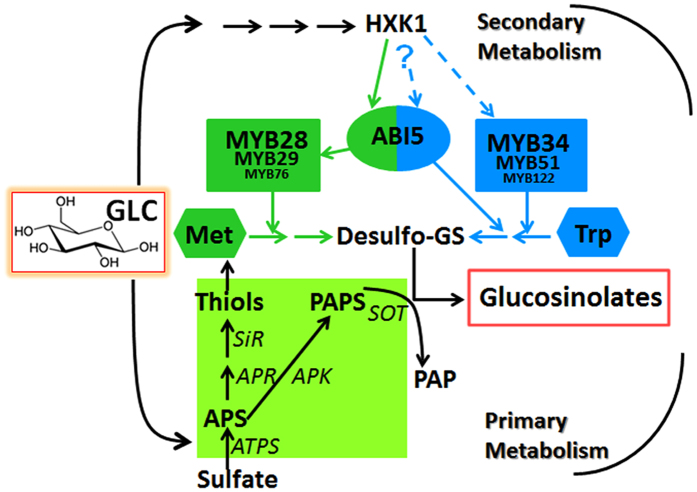
The effect of glucose as a signaling molecule on induction of aliphatic glucosinolate biosynthesis was reported in our former study. Here, we further investigated the regulatory mechanism of indolic glucosinolate biosynthesis by glucose in Arabidopsis. Glucose exerted a positive influence on indolic glucosinolate biosynthesis, which was demonstrated by induced accumulation of indolic glucosinolates and enhanced expression of related genes upon glucose treatment. Genetic analysis revealed that MYB34 and MYB51 were crucial in maintaining the basal indolic glucosinolate accumulation, with MYB34 being pivotal in response to glucose signaling. The increased accumulation of indolic glucosinolates and mRNA levels of MYB34, MYB51, and MYB122 caused by glucose were inhibited in the gin2-1 mutant, suggesting an important role of HXK1 in glucose-mediated induction of indolic glucosinolate biosynthesis. In contrast to what was known on the function of ABI5 in glucose-mediated aliphatic glucosinolate biosynthesis, ABI5 was not required for glucose-induced indolic glucosinolate accumulation. In addition, our results also indicated that glucose-induced glucosinolate accumulation was due to enhanced sulfur assimilation instead of directed sulfur partitioning into glucosinolate biosynthesis. Thus, our data provide new insights into molecular mechanisms underlying glucose-regulated glucosinolate biosynthesis.
Figures
Similar articles
-
MYB34, MYB51, and MYB122 distinctly regulate indolic glucosinolate biosynthesis in Arabidopsis thaliana.Mol Plant. 2014 May;7(5):814-28. doi: 10.1093/mp/ssu004. Epub 2014 Jan 15. Mol Plant. 2014. PMID: 24431192
-
The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana.Plant J. 2007 Jun;50(5):886-901. doi: 10.1111/j.1365-313X.2007.03099.x. Epub 2007 Apr 25. Plant J. 2007. PMID: 17461791
-
Glucose signalling positively regulates aliphatic glucosinolate biosynthesis.J Exp Bot. 2013 Feb;64(4):1097-109. doi: 10.1093/jxb/ers399. Epub 2013 Jan 16. J Exp Bot. 2013. PMID: 23329848 Free PMC article.
-
A Comprehensive Gene Inventory for Glucosinolate Biosynthetic Pathway in Arabidopsis thaliana.J Agric Food Chem. 2020 Jul 15;68(28):7281-7297. doi: 10.1021/acs.jafc.0c01916. Epub 2020 Jul 1. J Agric Food Chem. 2020. PMID: 32551569 Review.
-
The effect of sulfur nutrition on plant glucosinolate content: physiology and molecular mechanisms.Plant Biol (Stuttg). 2007 Sep;9(5):573-81. doi: 10.1055/s-2007-965431. Plant Biol (Stuttg). 2007. PMID: 17853357 Review.
Cited by
-
The SLIM1 transcription factor affects sugar signaling during sulfur deficiency in Arabidopsis.J Exp Bot. 2022 Dec 8;73(22):7362-7379. doi: 10.1093/jxb/erac371. J Exp Bot. 2022. PMID: 36099003 Free PMC article.
-
Low-temperature storage after harvest retards the deterioration in the sensory quality, health-promoting compounds, and antioxidant capacity of baby mustard.RSC Adv. 2020 Oct 5;10(60):36495-36503. doi: 10.1039/d0ra07177c. eCollection 2020 Oct 1. RSC Adv. 2020. PMID: 35517928 Free PMC article.
-
Blue Light Enhances Health-Promoting Sulforaphane Accumulation in Broccoli (Brassica oleracea var. italica) Sprouts through Inhibiting Salicylic Acid Synthesis.Plants (Basel). 2023 Sep 1;12(17):3151. doi: 10.3390/plants12173151. Plants (Basel). 2023. PMID: 37687397 Free PMC article.
-
Enhancing health-promoting isothiocyanates in Chinese kale sprouts via manipulating BoESP.Hortic Res. 2023 Feb 21;10(4):uhad029. doi: 10.1093/hr/uhad029. eCollection 2023 Apr. Hortic Res. 2023. PMID: 37090092 Free PMC article.
-
CRISPR/Cas9-Mediated Knockout of HOS1 Reveals Its Role in the Regulation of Secondary Metabolism in Arabidopsis thaliana.Plants (Basel). 2021 Jan 6;10(1):104. doi: 10.3390/plants10010104. Plants (Basel). 2021. PMID: 33419060 Free PMC article.
References
-
- Grubb C. D. & Abel S. Glucosinolate metabolism and its control. Trends Plant Sci 11, 89–100 (2006). - PubMed
-
- Kim J. H., Lee B. W., Schroeder F. C. & Jander G. Identification of indole glucosinolate breakdown products with antifeedant effects on Myzus persicae (green peach aphid). Plant J 54, 1015–1026 (2008). - PubMed
-
- Bednarek P. et al.. A Glucosinolate Metabolism Pathway in Living Plant Cells Mediates Broad-Spectrum Antifungal Defense. Science 323, 101–106 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
