

Stereospecific induction of apoptosis in tumor cells via endogenous C16-ceramide and distinct transcripts
- PMID: 27551447
- PMCID: PMC4979478
- DOI: 10.1038/cddiscovery.2015.13
Stereospecific induction of apoptosis in tumor cells via endogenous C16-ceramide and distinct transcripts
Abstract
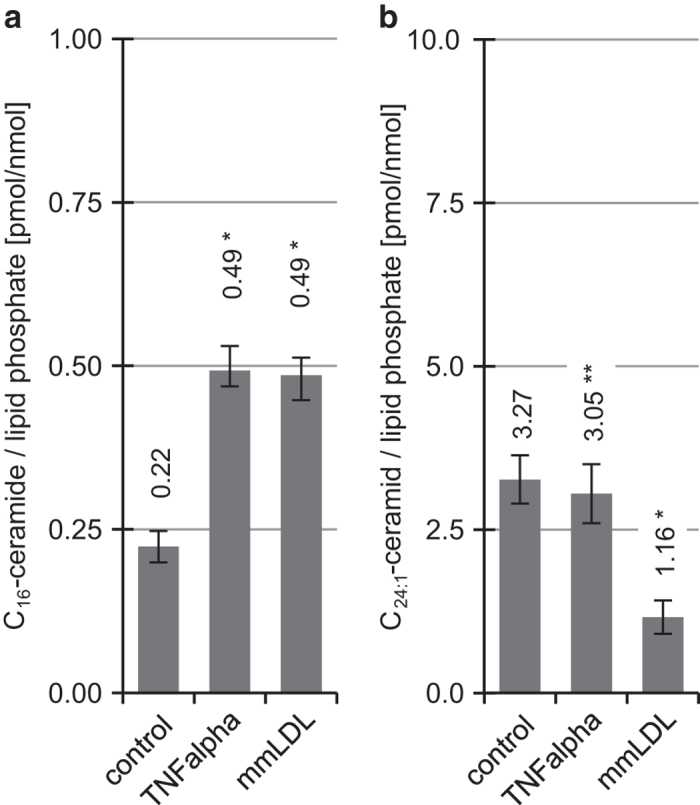
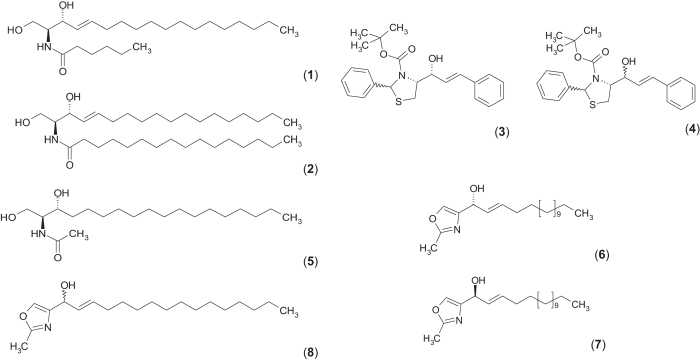
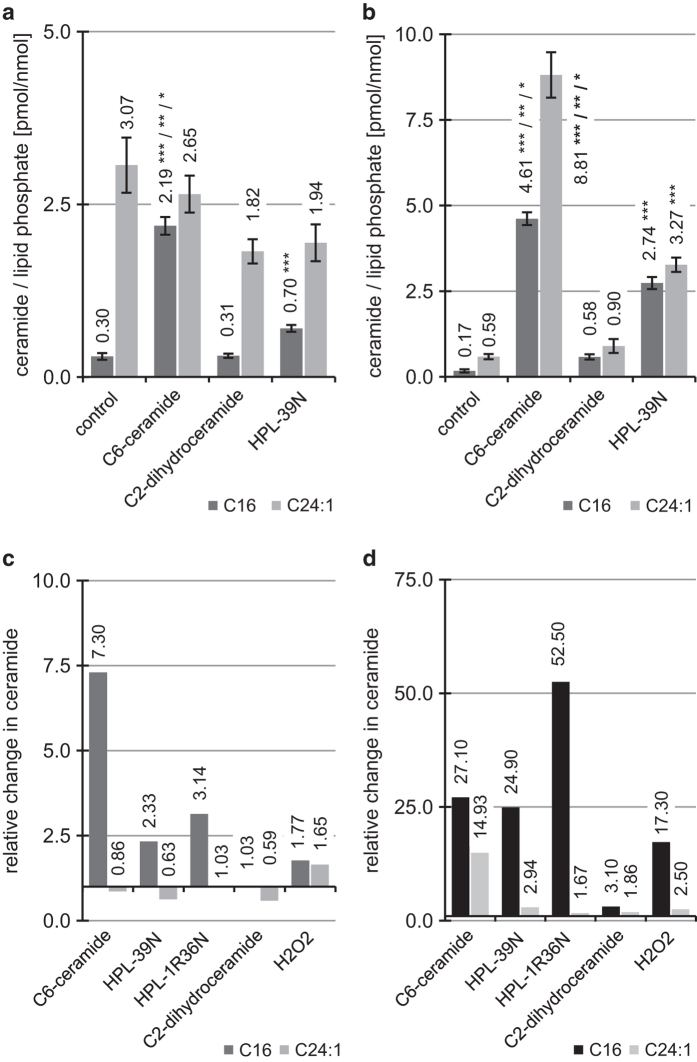
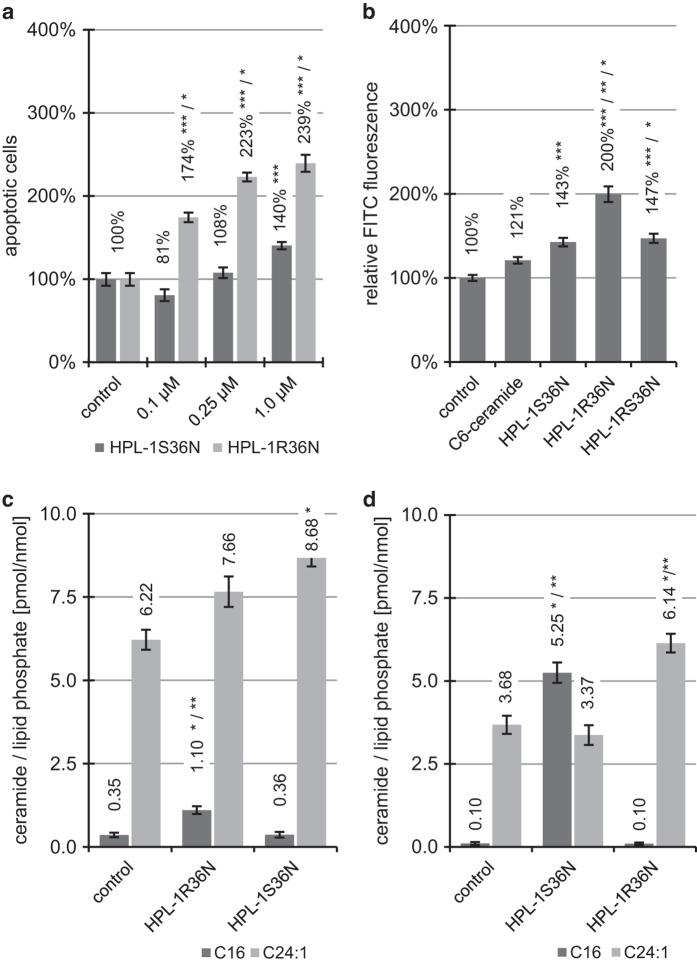
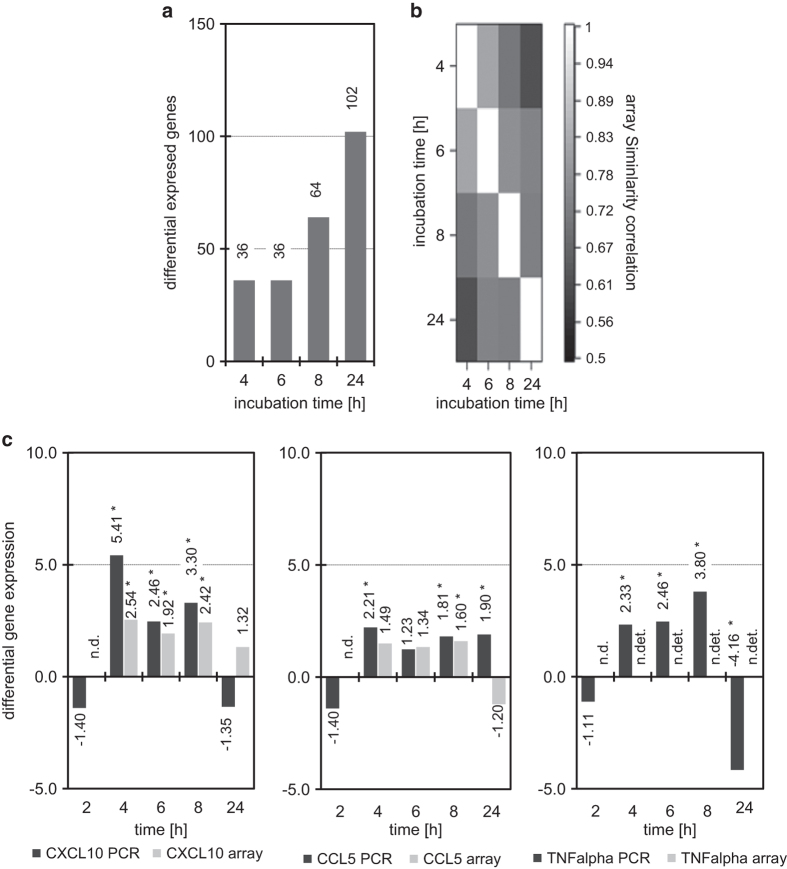
Concentration and distribution of individual endogenous ceramide species is crucial for apoptosis induction in response to various stimuli. Exogenous ceramide analogs induce apoptosis and can in turn modify the composition/concentrations of endogenous ceramide species and associated signaling. In this study, we show here that the elevation of endogenous C16-ceramide levels is a common feature of several known apoptosis-inducing triggers like mmLDL, TNF-alpha, H2O2 and exogenous C6-ceramide. Vice versa apoptosis requires elevation of endogenous C16-ceramide levels in cells. Enantiomers of a synthetic ceramide analog HPL-1RS36N have been developed as probes and vary in their capacity to inducing apoptosis in macrophages and HT-29 cells. Apoptosis induction by the two synthetic ceramide analogs HPL-39N and HPL-1R36N correlates with generation of cellular C16-ceramide concentration. In contrast to the S-enantiomer HPL-1S36N, the R-enantiomer HPL-1R36N shows significant effects on the expression of distinct genes known to be involved in cell cycle, cell growth and cell death (CXCL10, CCL5 and TNF-alpha), similarly on apoptosis induction. Enantioselective effects on transcription induced by metabolically stable synthetic probes provide clues on molecular mechanisms of ceramide-induced signaling, as well as leads for future anti-cancer agents.
Figures
Similar articles
-
The ceramide analog, B13, induces apoptosis in prostate cancer cell lines and inhibits tumor growth in prostate cancer xenografts.Prostate. 2004 Mar 1;58(4):382-93. doi: 10.1002/pros.10350. Prostate. 2004. PMID: 14968439
-
Tumor necrosis factor-alpha sensitizes prostate cancer cells to gamma-irradiation-induced apoptosis.Cancer Res. 1999 Apr 1;59(7):1606-14. Cancer Res. 1999. PMID: 10197636
-
Mechanisms involved in exogenous C2- and C6-ceramide-induced cancer cell toxicity.Biochem Pharmacol. 2003 May 15;65(10):1633-42. doi: 10.1016/s0006-2952(03)00125-4. Biochem Pharmacol. 2003. PMID: 12754099
-
Mechanisms of Ceramide-Dependent Cancer Cell Death.Adv Cancer Res. 2018;140:1-25. doi: 10.1016/bs.acr.2018.04.007. Adv Cancer Res. 2018. PMID: 30060806 Review.
-
Ceramide regulates cellular homeostasis via diverse stress signaling pathways.Leukemia. 2001 Aug;15(8):1153-60. doi: 10.1038/sj.leu.2402197. Leukemia. 2001. PMID: 11480555 Review.
Cited by
-
Ceramide Synthase 6 Maximizes p53 Function to Prevent Progeny Formation from Polyploid Giant Cancer Cells.Cancers (Basel). 2021 May 5;13(9):2212. doi: 10.3390/cancers13092212. Cancers (Basel). 2021. PMID: 34062962 Free PMC article.
-
A Novel Non-Psychoactive Fatty Acid from a Marine Snail, Conus inscriptus, Signals Cannabinoid Receptor 1 (CB1) to Accumulate Apoptotic C16:0 and C18:0 Ceramides in Teratocarcinoma Cell Line PA1.Molecules. 2024 Apr 11;29(8):1737. doi: 10.3390/molecules29081737. Molecules. 2024. PMID: 38675558 Free PMC article.
-
Derailed Ceramide Metabolism in Atopic Dermatitis (AD): A Causal Starting Point for a Personalized (Basic) Therapy.Int J Mol Sci. 2019 Aug 15;20(16):3967. doi: 10.3390/ijms20163967. Int J Mol Sci. 2019. PMID: 31443157 Free PMC article. Review.
-
C16:0 ceramide effect on melanoma malignant behavior and glycolysis depends on its intracellular or exogenous location.Am J Transl Res. 2020 Mar 15;12(3):1123-1135. eCollection 2020. Am J Transl Res. 2020. PMID: 32269739 Free PMC article.
-
C2-Ceramide-Induced Rb-Dominant Senescence-Like Phenotype Leads to Human Breast Cancer MCF-7 Escape from p53-Dependent Cell Death.Int J Mol Sci. 2019 Sep 2;20(17):4292. doi: 10.3390/ijms20174292. Int J Mol Sci. 2019. PMID: 31480728 Free PMC article.
References
-
- Van Brocklyn JR , Williams JB . The control of the balance between ceramide and sphingosine-1-phosphate by sphingosine kinase: oxidative stress and the seesaw of cell survival and death. Comp Biochem Physiol B Biochem Mol Biol 2012; 163: 26–36. - PubMed
-
- Saddoughi SA , Ogretmen B . Diverse functions of ceramide in cancer cell death and proliferation. Adv Cancer Res 2013; 117: 37–58. - PubMed
-
- Claus RA , Dorer MJ , Bunck AC , Deigner H-P . Inhibition of sphingomyelin hydrolysis: targeting the lipid mediator ceramide as a key regulator of cellular fate. Curr Med Chem 2009; 16: 1978–2000. - PubMed
-
- Claus RA , Bunck AC , Bockmeyer CL , Brunkhorst FM , Lösche W , Kinscherf R et al. Role of increased sphingomyelinase activity in apoptosis and organ failure of patients with severe sepsis. FASEB J 2005; 19: 1719–1721. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
