

Identification of RNA Binding Proteins Associated with Dengue Virus RNA in Infected Cells Reveals Temporally Distinct Host Factor Requirements
- PMID: 27556644
- PMCID: PMC4996428
- DOI: 10.1371/journal.pntd.0004921
Identification of RNA Binding Proteins Associated with Dengue Virus RNA in Infected Cells Reveals Temporally Distinct Host Factor Requirements
Abstract
Background: There are currently no vaccines or antivirals available for dengue virus infection, which can cause dengue hemorrhagic fever and death. A better understanding of the host pathogen interaction is required to develop effective therapies to treat DENV. In particular, very little is known about how cellular RNA binding proteins interact with viral RNAs. RNAs within cells are not naked; rather they are coated with proteins that affect localization, stability, translation and (for viruses) replication.
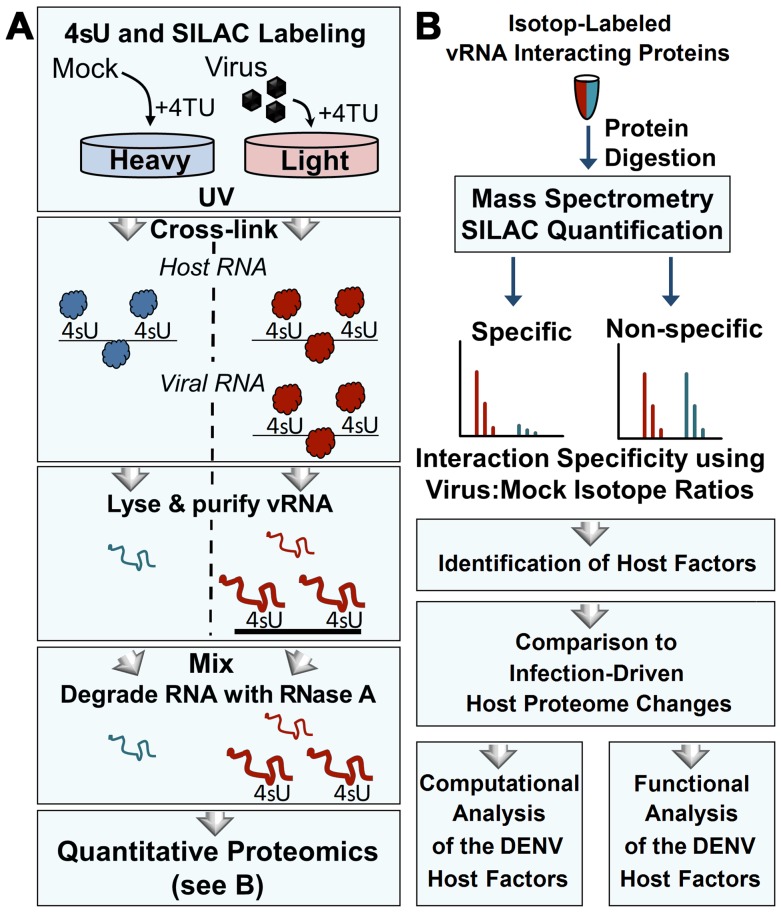
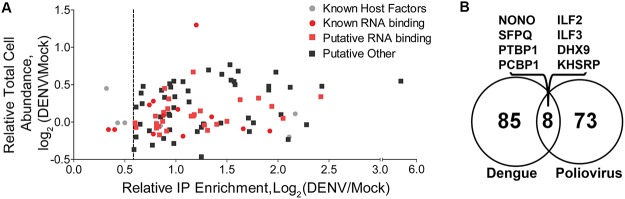
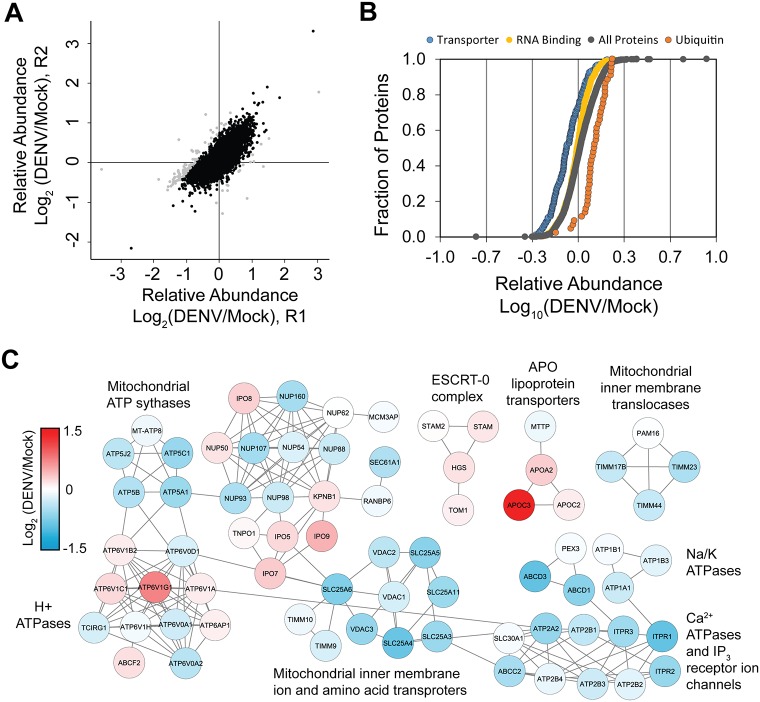
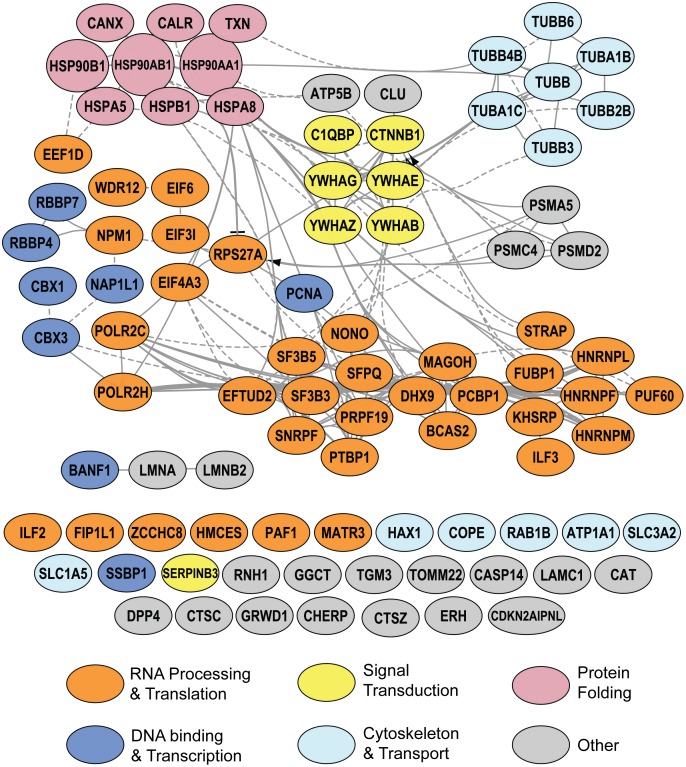
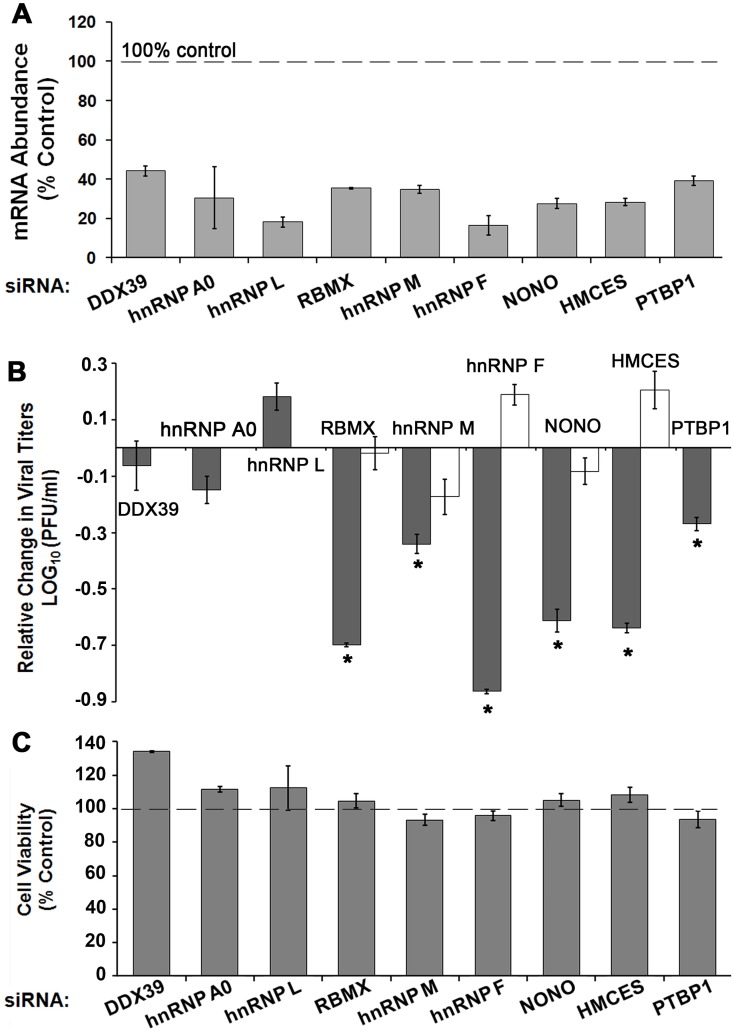
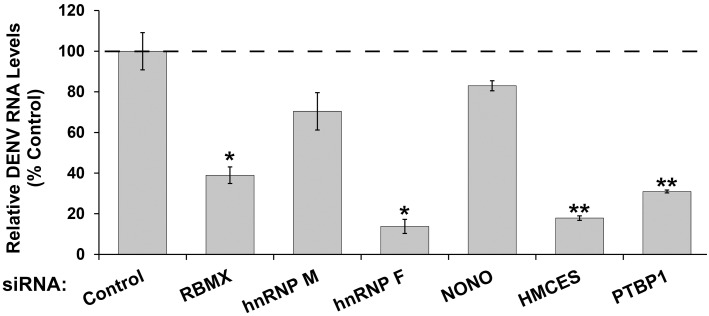
Methodology/principal findings: Seventy-nine novel RNA binding proteins for dengue virus (DENV) were identified by cross-linking proteins to dengue viral RNA during a live infection in human cells. These cellular proteins were specific and distinct from those previously identified for poliovirus, suggesting a specialized role for these factors in DENV amplification. Knockdown of these proteins demonstrated their function as viral host factors, with evidence for some factors acting early, while others late in infection. Their requirement by DENV for efficient amplification is likely specific, since protein knockdown did not impair the cell fitness for viral amplification of an unrelated virus. The protein abundances of these host factors were not significantly altered during DENV infection, suggesting their interaction with DENV RNA was due to specific recruitment mechanisms. However, at the global proteome level, DENV altered the abundances of proteins in particular classes, including transporter proteins, which were down regulated, and proteins in the ubiquitin proteasome pathway, which were up regulated.
Conclusions/significance: The method for identification of host factors described here is robust and broadly applicable to all RNA viruses, providing an avenue to determine the conserved or distinct mechanisms through which diverse viruses manage the viral RNA within cells. This study significantly increases the number of cellular factors known to interact with DENV and reveals how DENV modulates and usurps cellular proteins for efficient amplification.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
