

Functional assessment of SLC4A11, an integral membrane protein mutated in corneal dystrophies
- PMID: 27558157
- PMCID: PMC5130586
- DOI: 10.1152/ajpcell.00078.2016
Functional assessment of SLC4A11, an integral membrane protein mutated in corneal dystrophies
Abstract
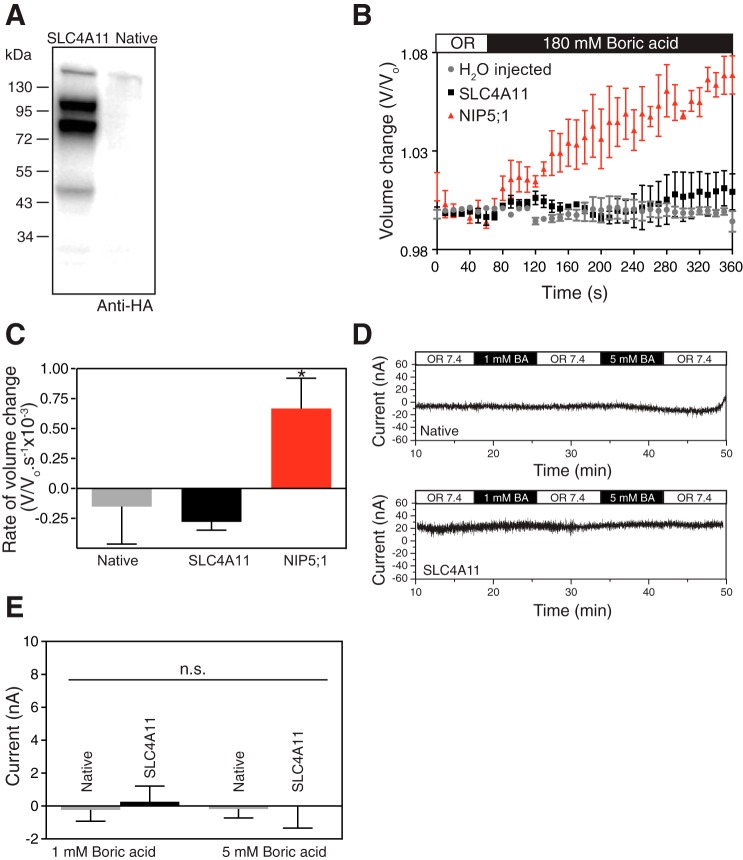
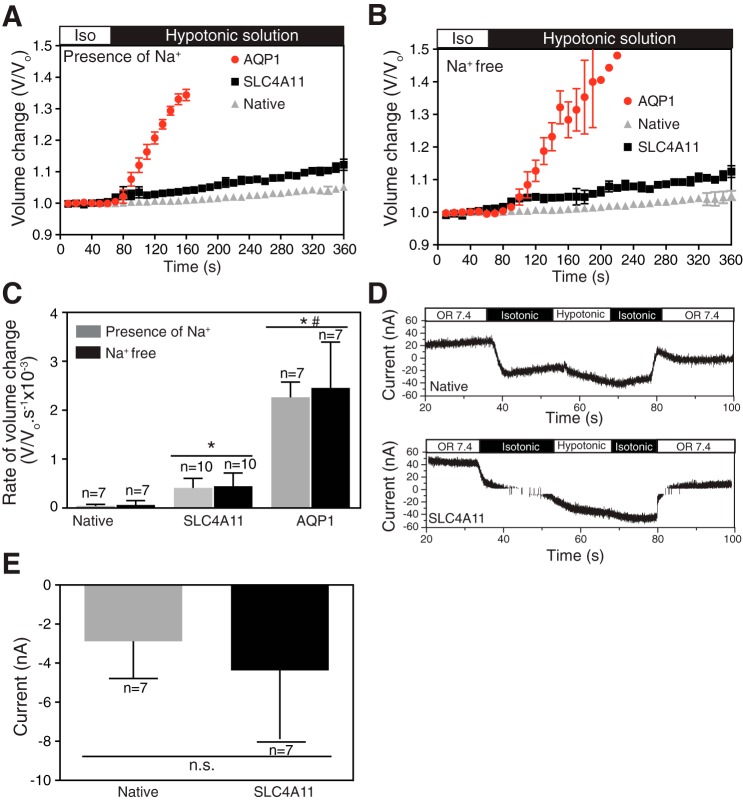
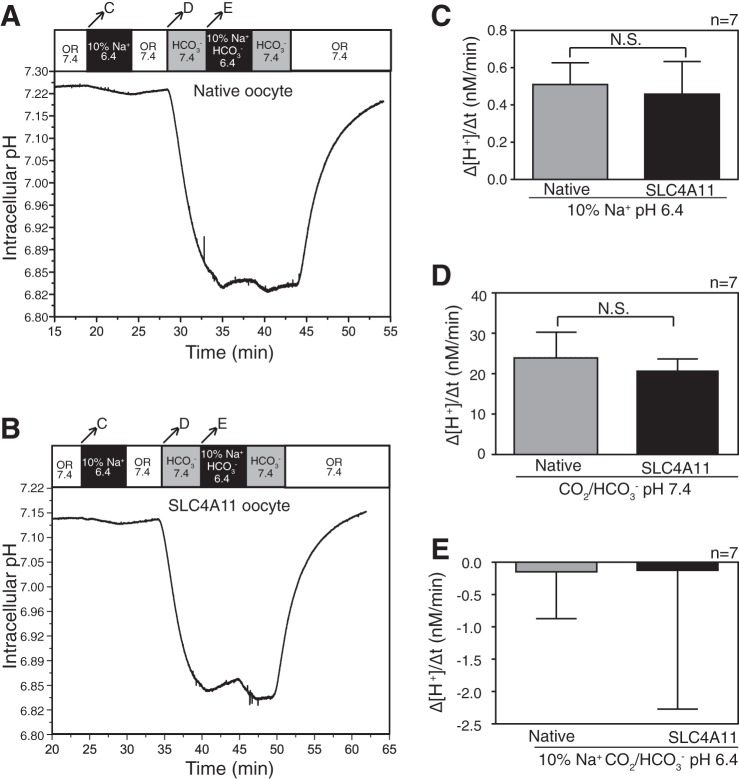
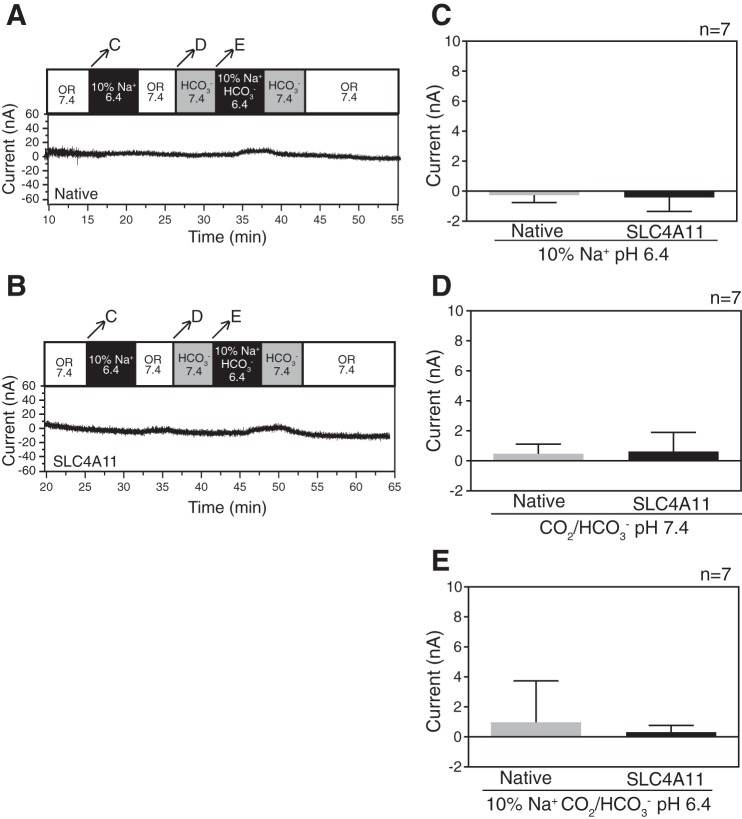
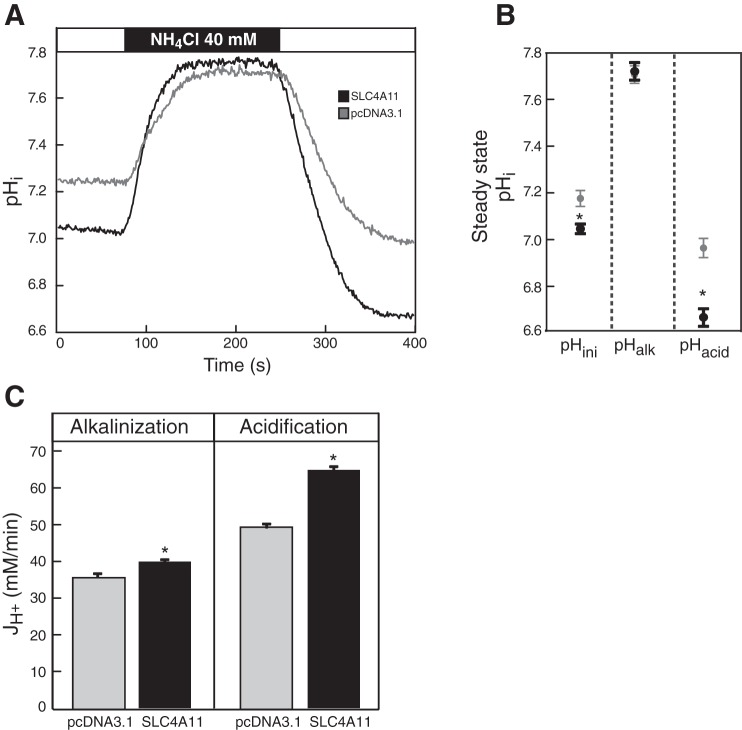
SLC4A11, a member of the SLC4 family of bicarbonate transporters, is a widely expressed integral membrane protein, abundant in kidney and cornea. Mutations of SLC4A11 cause some cases of the blinding corneal dystrophies, congenital hereditary endothelial dystrophy, and Fuchs endothelial corneal dystrophy. These diseases are marked by fluid accumulation in the corneal stroma, secondary to defective fluid reabsorption by the corneal endothelium. The role of SLC4A11 in these corneal dystrophies is not firmly established, as SLC4A11 function remains unclear. To clarify the normal function(s) of SLC4A11, we characterized the protein following expression in the simple, low-background expression system Xenopus laevis oocytes. Since plant and fungal SLC4A11 orthologs transport borate, we measured cell swelling associated with accumulation of solute borate. The plant water/borate transporter NIP5;1 manifested borate transport, whereas human SLC4A11 did not. SLC4A11 supported osmotically driven water accumulation that was electroneutral and Na+ independent. Studies in oocytes and HEK293 cells could not detect Na+-coupled HCO3- transport or Cl-/HCO3- exchange by SLC4A11. SLC4A11 mediated electroneutral NH3 transport in oocytes. Voltage-dependent OH- or H+ movement was not measurable in SLC4A11-expressing oocytes, but SLC4A11-expressing HEK293 cells manifested low-level cytosolic acidification at baseline. In mammalian cells, but not oocytes, OH-/H+ conductance may arise when SLC4A11 activates another protein or itself is activated by another protein. These data argue against a role of human SLC4A11 in bicarbonate or borate transport. This work provides additional support for water and ammonia transport by SLC4A11. When expressed in oocytes, SLC4A11 transported NH3, not NH3/H.
Keywords: SLC4A11; ammonia; corneal dystrophy; endothelial cell; water flux.
Copyright © 2016 the American Physiological Society.
Figures
References
-
- Aldahmesh MA, Khan AO, Meyer BF, Alkuraya FS. Mutational spectrum of SLC4A11 in autosomal recessive CHED in Saudi Arabia. Invest Ophthalmol Vis Sci 50: 4142–4145, 2009. - PubMed
-
- Aldave AJ, Yellore VS, Bourla N, Momi RS, Khan MA, Salem AK, Rayner SA, Glasgow BJ, Kurtz I. Autosomal recessive CHED associated with novel compound heterozygous mutations in SLC4A11. Cornea 26: 896–900, 2007. - PubMed
-
- Alka K, Casey JR. Bicarbonate transport in health and disease. IUBMB Life 66: 596–615, 2014. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources