

A Quantitative Characterization of Nucleoplasmin/Histone Complexes Reveals Chaperone Versatility
- PMID: 27558753
- PMCID: PMC4997359
- DOI: 10.1038/srep32114
A Quantitative Characterization of Nucleoplasmin/Histone Complexes Reveals Chaperone Versatility
Abstract
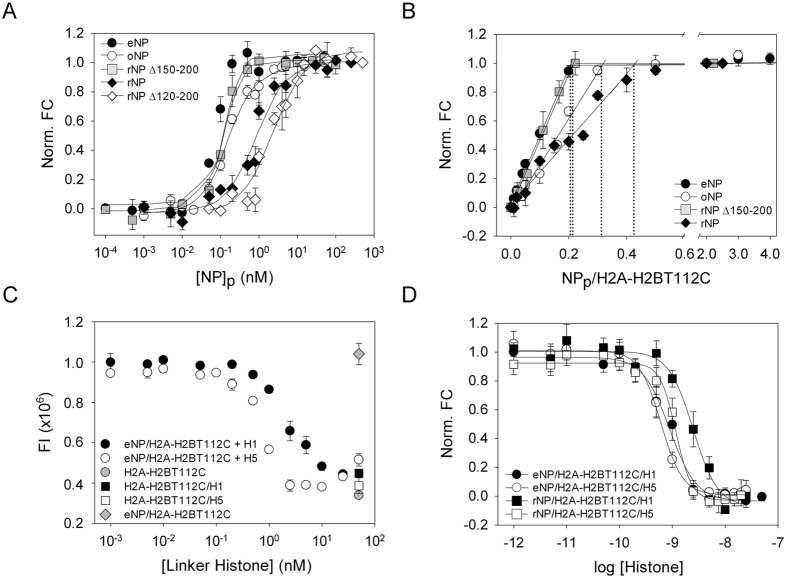
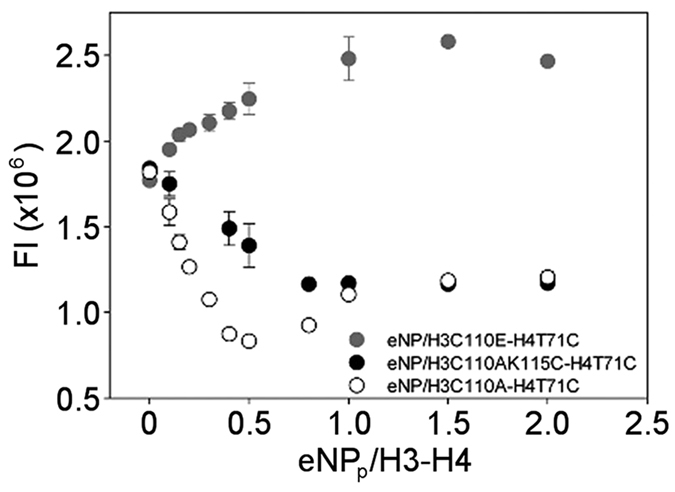
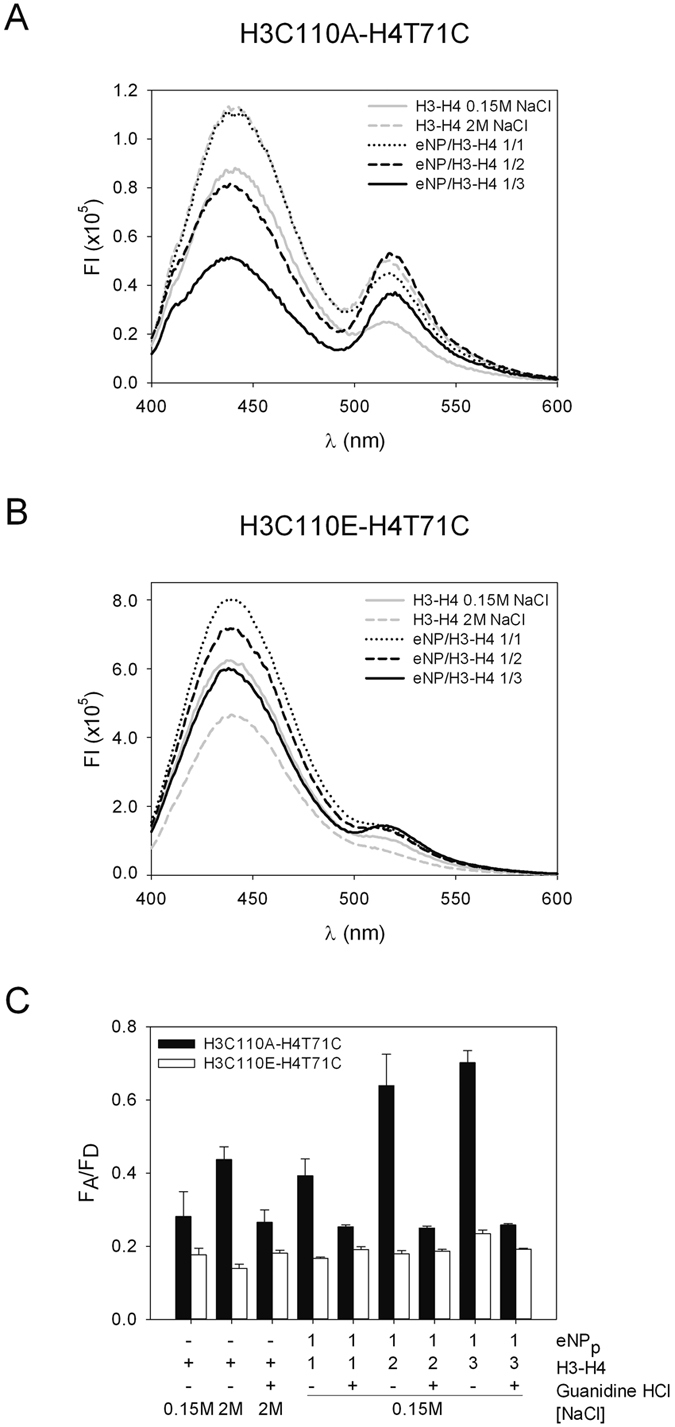
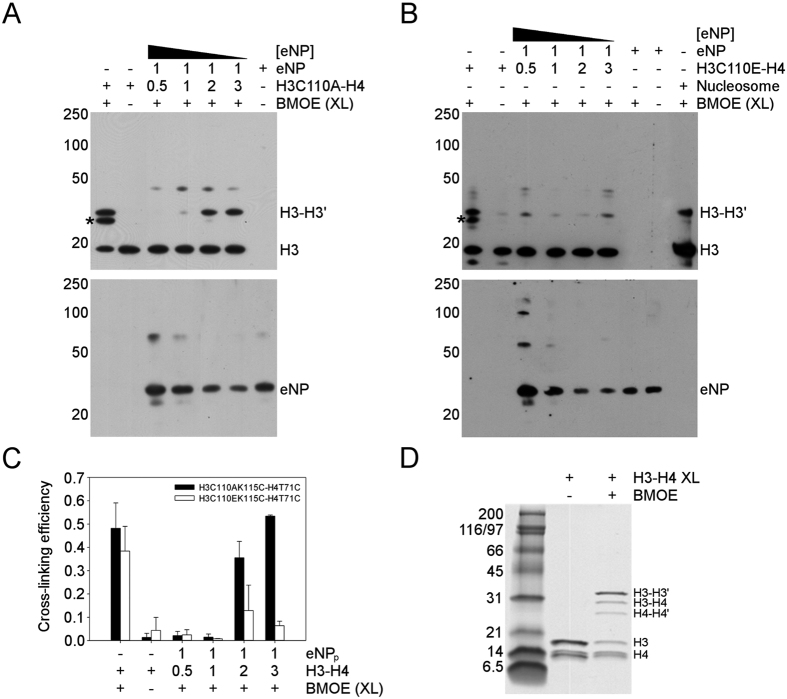
Nucleoplasmin (NP) is an abundant histone chaperone in vertebrate oocytes and embryos involved in storing and releasing maternal histones to establish and maintain the zygotic epigenome. NP has been considered a H2A-H2B histone chaperone, and recently it has been shown that it can also interact with H3-H4. However, its interaction with different types of histones has not been quantitatively studied so far. We show here that NP binds H2A-H2B, H3-H4 and linker histones with Kd values in the subnanomolar range, forming different complexes. Post-translational modifications of NP regulate exposure of the polyGlu tract at the disordered distal face of the protein and induce an increase in chaperone affinity for all histones. The relative affinity of NP for H2A-H2B and linker histones and the fact that they interact with the distal face of the chaperone could explain their competition for chaperone binding, a relevant process in NP-mediated sperm chromatin remodelling during fertilization. Our data show that NP binds H3-H4 tetramers in a nucleosomal conformation and dimers, transferring them to DNA to form disomes and tetrasomes. This finding might be relevant to elucidate the role of NP in chromatin disassembly and assembly during replication and transcription.
Figures



Similar articles
-
Structural insights into the ability of nucleoplasmin to assemble and chaperone histone octamers for DNA deposition.Sci Rep. 2019 Jul 1;9(1):9487. doi: 10.1038/s41598-019-45726-7. Sci Rep. 2019. PMID: 31263230 Free PMC article.
-
A mechanism for histone chaperoning activity of nucleoplasmin: thermodynamic and structural models.J Mol Biol. 2009 Oct 23;393(2):448-63. doi: 10.1016/j.jmb.2009.08.005. Epub 2009 Aug 13. J Mol Biol. 2009. PMID: 19683001
-
The intrinsically disordered distal face of nucleoplasmin recognizes distinct oligomerization states of histones.Nucleic Acids Res. 2014 Jan;42(2):1311-25. doi: 10.1093/nar/gkt899. Epub 2013 Oct 10. Nucleic Acids Res. 2014. PMID: 24121686 Free PMC article.
-
The role of nucleoplasmin in chromatin assembly and disassembly.Philos Trans R Soc Lond B Biol Sci. 1993 Mar 29;339(1289):263-9; discussion 268-9. doi: 10.1098/rstb.1993.0024. Philos Trans R Soc Lond B Biol Sci. 1993. PMID: 8098530 Review.
-
Vertebrate nucleoplasmin and NASP: egg histone storage proteins with multiple chaperone activities.FASEB J. 2012 Dec;26(12):4788-804. doi: 10.1096/fj.12-216663. Epub 2012 Sep 11. FASEB J. 2012. PMID: 22968912 Review.
Cited by
-
Structure-function studies of a nucleoplasmin isoform from Plasmodium falciparum.J Biol Chem. 2025 Apr;301(4):108379. doi: 10.1016/j.jbc.2025.108379. Epub 2025 Mar 4. J Biol Chem. 2025. PMID: 40049416 Free PMC article.
-
Double maternal-effect: duplicated nucleoplasmin 2 genes, npm2a and npm2b, with essential but distinct functions are shared by fish and tetrapods.BMC Evol Biol. 2018 Nov 12;18(1):167. doi: 10.1186/s12862-018-1281-3. BMC Evol Biol. 2018. PMID: 30419815 Free PMC article.
-
Plant-specific HDT family histone deacetylases are nucleoplasmins.Plant Cell. 2022 Nov 29;34(12):4760-4777. doi: 10.1093/plcell/koac275. Plant Cell. 2022. PMID: 36069647 Free PMC article.
-
Structural insights into the ability of nucleoplasmin to assemble and chaperone histone octamers for DNA deposition.Sci Rep. 2019 Jul 1;9(1):9487. doi: 10.1038/s41598-019-45726-7. Sci Rep. 2019. PMID: 31263230 Free PMC article.
-
AtFKBP53: a chimeric histone chaperone with functional nucleoplasmin and PPIase domains.Nucleic Acids Res. 2020 Feb 20;48(3):1531-1550. doi: 10.1093/nar/gkz1153. Nucleic Acids Res. 2020. PMID: 31807785 Free PMC article.
References
-
- Leno G. H., Mills A. D., Philpott A. & Laskey R. A. Hyperphosphorylation of nucleoplasmin facilitates Xenopus sperm decondensation at fertilization. J Biol Chem 271, 7253–7256 (1996). - PubMed
-
- Leno G. H. Cell-free systems to study chromatin remodeling. Methods Cell Biol 53, 497–515 (1998). - PubMed
-
- Tyler J. K. Chromatin assembly. Cooperation between histone chaperones and ATP-dependent nucleosome remodeling machines. Eur J Biochem/FEBS 269, 2268–2274 (2002). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
