

Glycoproteomics Reveals Decorin Peptides With Anti-Myostatin Activity in Human Atrial Fibrillation
- PMID: 27559042
- PMCID: PMC5081096
- DOI: 10.1161/CIRCULATIONAHA.115.016423
Glycoproteomics Reveals Decorin Peptides With Anti-Myostatin Activity in Human Atrial Fibrillation
Abstract
Background: Myocardial fibrosis is a feature of many cardiac diseases. We used proteomics to profile glycoproteins in the human cardiac extracellular matrix (ECM).
Methods: Atrial specimens were analyzed by mass spectrometry after extraction of ECM proteins and enrichment for glycoproteins or glycopeptides.
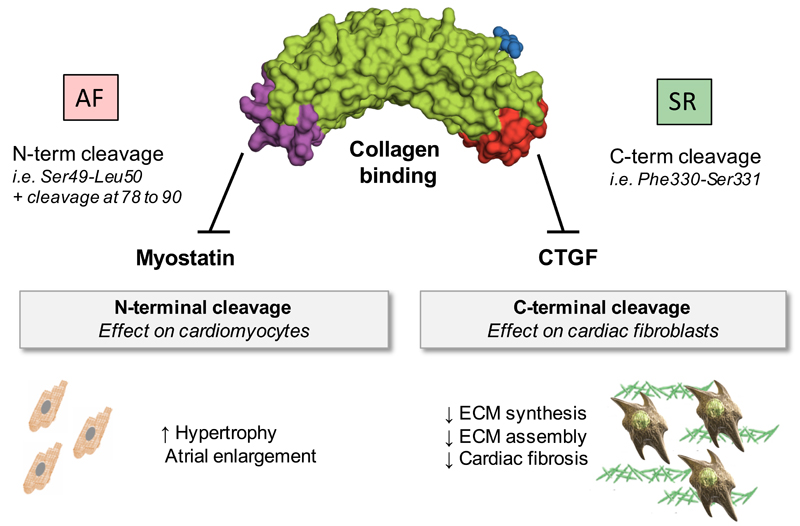
Results: ECM-related glycoproteins were identified in left and right atrial appendages from the same patients. Several known glycosylation sites were confirmed. In addition, putative and novel glycosylation sites were detected. On enrichment for glycoproteins, peptides of the small leucine-rich proteoglycan decorin were identified consistently in the flowthrough. Of all ECM proteins identified, decorin was found to be the most fragmented. Within its protein core, 18 different cleavage sites were identified. In contrast, less cleavage was observed for biglycan, the most closely related proteoglycan. Decorin processing differed between human ventricles and atria and was altered in disease. The C-terminus of decorin, important for the interaction with connective tissue growth factor, was detected predominantly in ventricles in comparison with atria. In contrast, atrial appendages from patients in persistent atrial fibrillation had greater levels of full-length decorin but also harbored a cleavage site that was not found in atrial appendages from patients in sinus rhythm. This cleavage site preceded the N-terminal domain of decorin that controls muscle growth by altering the binding capacity for myostatin. Myostatin expression was decreased in atrial appendages of patients with persistent atrial fibrillation and hearts of decorin null mice. A synthetic peptide corresponding to this decorin region dose-dependently inhibited the response to myostatin in cardiomyocytes and in perfused mouse hearts.
Conclusions: This proteomics study is the first to analyze the human cardiac ECM. Novel processed forms of decorin protein core, uncovered in human atrial appendages, can regulate the local bioavailability of antihypertrophic and profibrotic growth factors.
Keywords: atrial fibrillation; cardiovascular diseases; extracellular matrix; mass spectrometry; proteomics.
© 2016 American Heart Association, Inc.
Figures






References
-
- Dawson K, Wakili R, Ordög B, Clauss S, Chen Y, Iwasaki Y, Voigt N, Qi XY, Sinner MF, Dobrev D, Kääb S, et al. MicroRNA29: a mechanistic contributor and potential biomarker in atrial fibrillation. Circulation. 2013;127:1466–1475. - PubMed
-
- Mayr M, Yusuf S, Weir G, Chung YL, Mayr U, Yin X, Ladroue C, Madhu B, Roberts N, De Souza A, Fredericks S, et al. Combined metabolomic and proteomic analysis of human atrial fibrillation. J Am Coll Cardiol. 2008;51:585–594. - PubMed
-
- Barallobre-Barreiro J, Didangelos A, Schoendube FA, Drozdov I, Yin X, Fernández-Caggiano M, Willeit P, Puntmann VO, Aldama-López G, Shah AM, Doménech N, et al. Proteomics analysis of cardiac extracellular matrix remodeling in a porcine model of ischemia/reperfusion injury. Circulation. 2012;125:789–802. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
