

Neuromuscular NMDA Receptors Modulate Developmental Synapse Elimination
- PMID: 27559162
- PMCID: PMC6601906
- DOI: 10.1523/JNEUROSCI.1181-16.2016
Neuromuscular NMDA Receptors Modulate Developmental Synapse Elimination
Erratum in
-
Correction: Personius et al., "Neuromuscular NMDA Receptors Modulate Developmental Synapse Elimination".J Neurosci. 2018 Nov 7;38(45):9801. doi: 10.1523/JNEUROSCI.2401-18.2018. J Neurosci. 2018. PMID: 31329692 Free PMC article.
Abstract
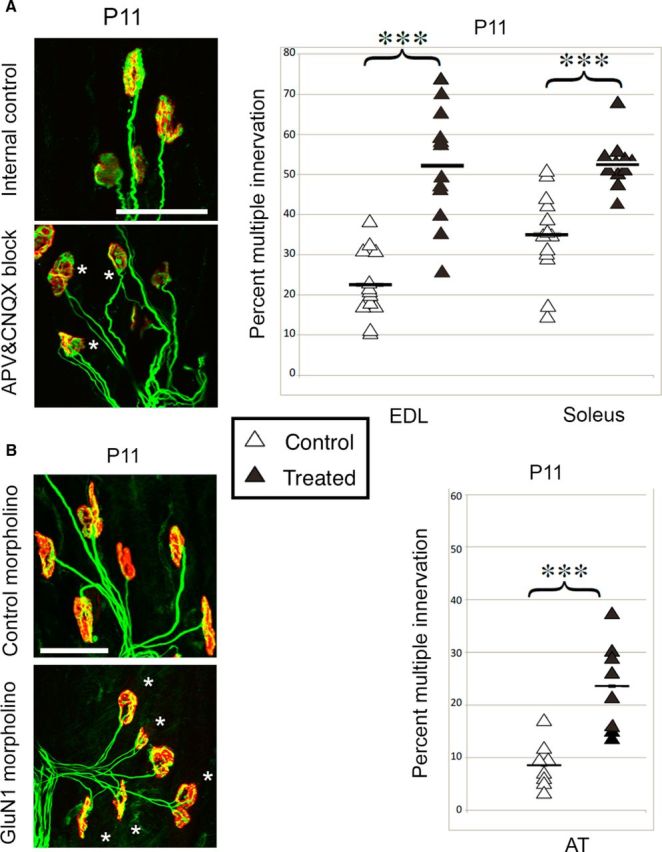
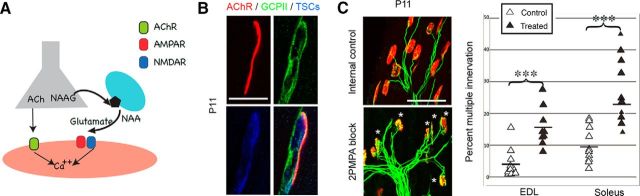
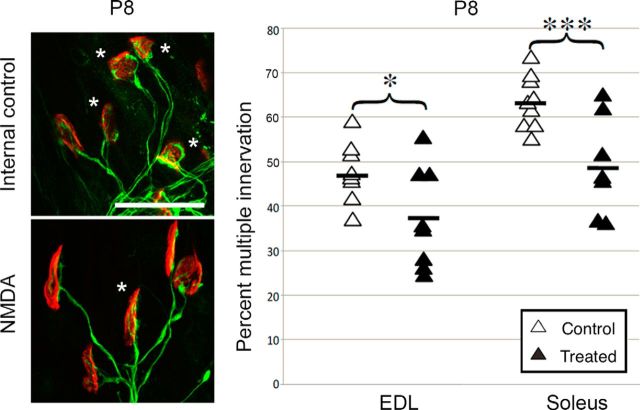
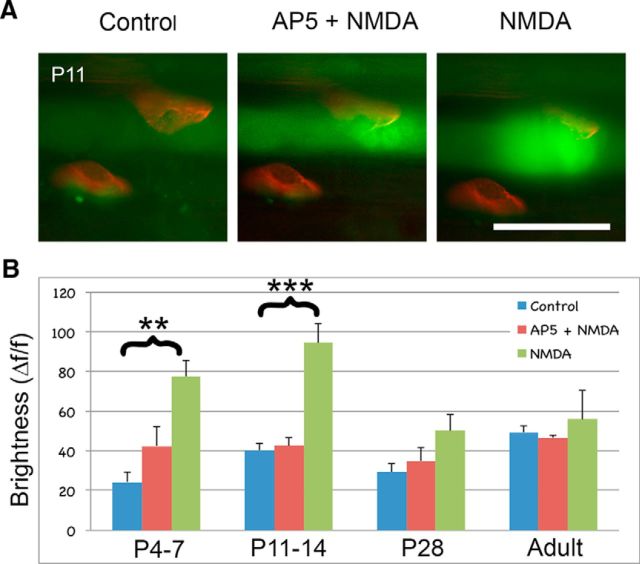
At birth, each mammalian skeletal muscle fiber is innervated by multiple motor neurons, but in a few weeks, all but one of those axons retracts (Redfern, 1970) and differential activity between inputs controls this phenomenon (Personius and Balice-Gordon, 2001; Sanes and Lichtman, 2001; Personius et al., 2007; Favero et al., 2012). Acetylcholine, the primary neuromuscular transmitter, has long been presumed to mediate this activity-dependent process (O'Brien et al., 1978), but glutamatergic transmission also occurs at the neuromuscular junction (Berger et al., 1995; Grozdanovic and Gossrau, 1998; Mays et al., 2009). To test the role of neuromuscular NMDA receptors, we assessed their contribution to muscle calcium fluxes in mice and tested whether they influence removal of excess innervation at the end plate. Developmental synapse pruning was slowed by reduction of NMDA receptor activation or expression and by reduction of glutamate production. Conversely, pruning is accelerated by application of exogenous NMDA. We also found that NMDA induced increased muscle calcium only during the first 2 postnatal weeks. Therefore, neuromuscular NMDA receptors play previously unsuspected roles in neuromuscular activity and synaptic pruning during development.
Significance statement: In normal adult muscle, each muscle fiber is innervated by a single axon, but at birth, fibers are multiply innervated. Elimination of excess connections requires neural activity; because the neuromuscular junction (NMJ) is a cholinergic synapse, acetylcholine has been assumed to be the critical mediator of activity. However, glutamate receptors are also expressed at the NMJ. We found that axon removal in mice is slowed by pharmacological and molecular manipulations that decrease signaling through neuromuscular NMDA receptors, whereas application of exogenous NMDA at the NMJ accelerates synapse elimination and increases muscle calcium levels during the first 2 postnatal weeks. Therefore, neuromuscular NMDA receptors play previously unsuspected roles in neuromuscular activity and elimination of excess synaptic input during development.
Keywords: competition; glutamate; neuromuscular junction; polyneuronal; synapse elimination.
Copyright © 2016 the authors 0270-6474/16/368783-07$15.00/0.
Figures
Similar articles
-
Developmental neuromuscular synapse elimination: Activity-dependence and potential downstream effector mechanisms.Neurosci Lett. 2020 Jan 23;718:134724. doi: 10.1016/j.neulet.2019.134724. Epub 2019 Dec 23. Neurosci Lett. 2020. PMID: 31877335 Free PMC article. Review.
-
Glutamatergic modulation of synaptic plasticity at a PNS vertebrate cholinergic synapse.Eur J Neurosci. 2003 Dec;18(12):3241-50. doi: 10.1111/j.1460-9568.2003.03028.x. Eur J Neurosci. 2003. PMID: 14686898
-
Decreased calcium influx into the neonatal rat motor nerve terminals can recruit additional neuromuscular junctions during the synapse elimination period.Neuroscience. 2002;110(1):147-54. doi: 10.1016/s0306-4522(01)00543-7. Neuroscience. 2002. PMID: 11882379
-
Presynaptic muscarinic acetylcholine autoreceptors (M1, M2 and M4 subtypes), adenosine receptors (A1 and A2A) and tropomyosin-related kinase B receptor (TrkB) modulate the developmental synapse elimination process at the neuromuscular junction.Mol Brain. 2016 Jun 23;9(1):67. doi: 10.1186/s13041-016-0248-9. Mol Brain. 2016. PMID: 27339059 Free PMC article.
-
Presynaptic Membrane Receptors Modulate ACh Release, Axonal Competition and Synapse Elimination during Neuromuscular Junction Development.Front Mol Neurosci. 2017 May 16;10:132. doi: 10.3389/fnmol.2017.00132. eCollection 2017. Front Mol Neurosci. 2017. PMID: 28559796 Free PMC article. Review.
Cited by
-
Developmental neuromuscular synapse elimination: Activity-dependence and potential downstream effector mechanisms.Neurosci Lett. 2020 Jan 23;718:134724. doi: 10.1016/j.neulet.2019.134724. Epub 2019 Dec 23. Neurosci Lett. 2020. PMID: 31877335 Free PMC article. Review.
-
NMDA receptors control development of somatosensory callosal axonal projections.Elife. 2021 Mar 4;10:e59612. doi: 10.7554/eLife.59612. Elife. 2021. PMID: 33661095 Free PMC article.
-
Skeletal muscle reprogramming enhances reinnervation after peripheral nerve injury.Nat Commun. 2024 Oct 25;15(1):9218. doi: 10.1038/s41467-024-53276-4. Nat Commun. 2024. PMID: 39455585 Free PMC article.
-
Homeostatic Plasticity of the Mammalian Neuromuscular Junction.Adv Neurobiol. 2022;28:111-130. doi: 10.1007/978-3-031-07167-6_5. Adv Neurobiol. 2022. PMID: 36066823
-
From Neural Tube Formation Through the Differentiation of Spinal Cord Neurons: Ion Channels in Action During Neural Development.Front Mol Neurosci. 2020 Apr 24;13:62. doi: 10.3389/fnmol.2020.00062. eCollection 2020. Front Mol Neurosci. 2020. PMID: 32390800 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources