

Minimally Mutated HIV-1 Broadly Neutralizing Antibodies to Guide Reductionist Vaccine Design
- PMID: 27560183
- PMCID: PMC4999182
- DOI: 10.1371/journal.ppat.1005815
Minimally Mutated HIV-1 Broadly Neutralizing Antibodies to Guide Reductionist Vaccine Design
Erratum in
-
Correction: Minimally Mutated HIV-1 Broadly Neutralizing Antibodies to Guide Reductionist Vaccine Design.PLoS Pathog. 2016 Sep 14;12(9):e1005905. doi: 10.1371/journal.ppat.1005905. eCollection 2016 Sep. PLoS Pathog. 2016. PMID: 27627763 Free PMC article.
Abstract
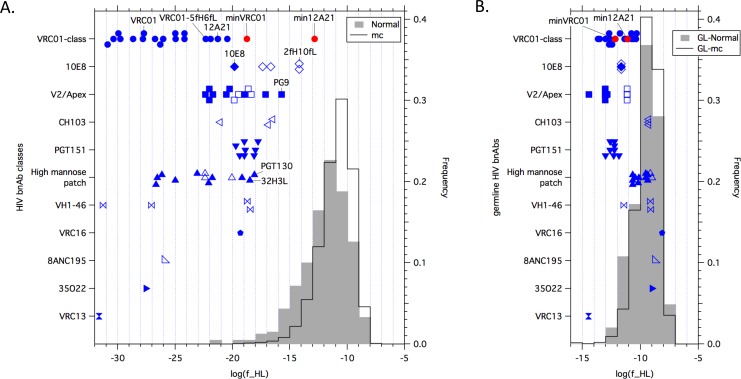
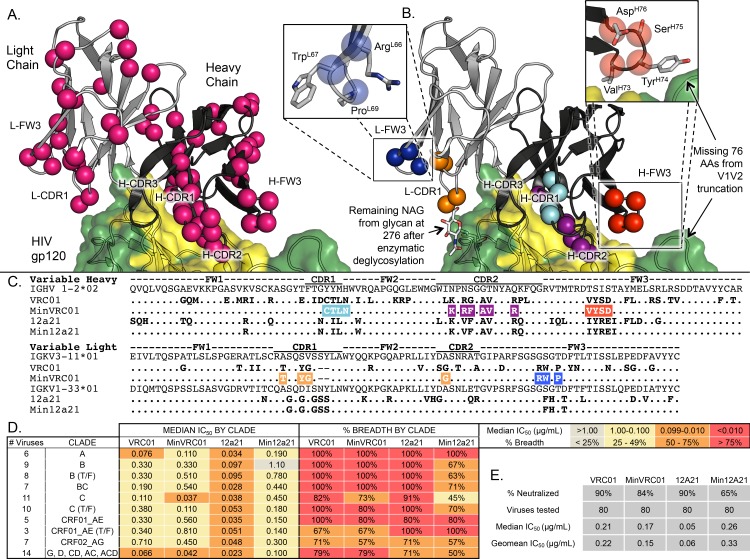
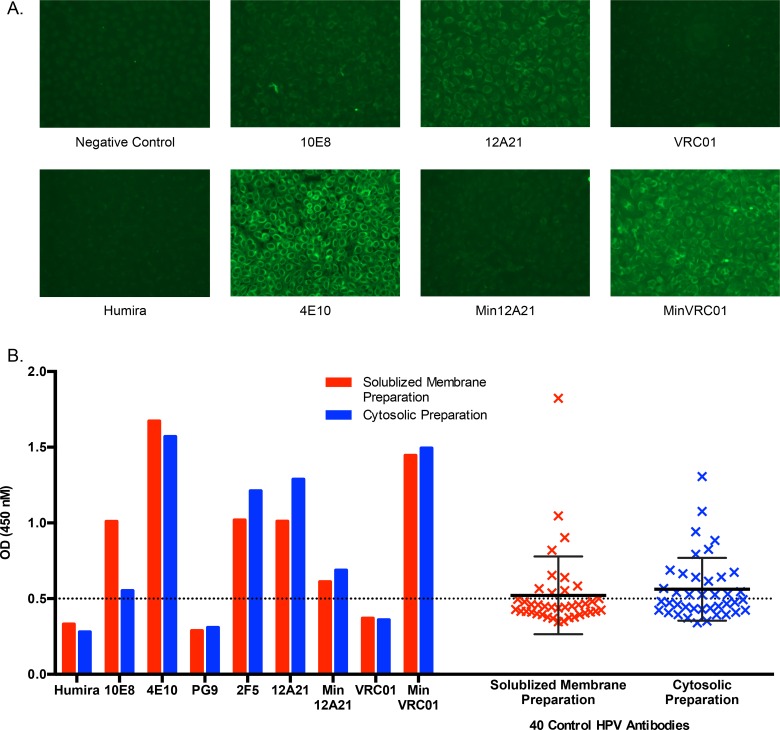
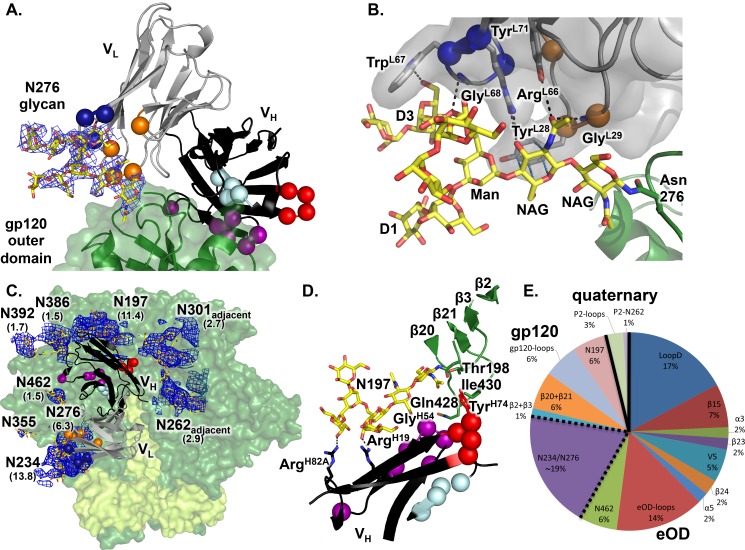
An optimal HIV vaccine should induce broadly neutralizing antibodies (bnAbs) that neutralize diverse viral strains and subtypes. However, potent bnAbs develop in only a small fraction of HIV-infected individuals, all contain rare features such as extensive mutation, insertions, deletions, and/or long complementarity-determining regions, and some are polyreactive, casting doubt on whether bnAbs to HIV can be reliably induced by vaccination. We engineered two potent VRC01-class bnAbs that minimized rare features. According to a quantitative features frequency analysis, the set of features for one of these minimally mutated bnAbs compared favorably with all 68 HIV bnAbs analyzed and was similar to antibodies elicited by common vaccines. This same minimally mutated bnAb lacked polyreactivity in four different assays. We then divided the minimal mutations into spatial clusters and dissected the epitope components interacting with those clusters, by mutational and crystallographic analyses coupled with neutralization assays. Finally, by synthesizing available data, we developed a working-concept boosting strategy to select the mutation clusters in a logical order following a germline-targeting prime. We have thus developed potent HIV bnAbs that may be more tractable vaccine goals compared to existing bnAbs, and we have proposed a strategy to elicit them. This reductionist approach to vaccine design, guided by antibody and antigen structure, could be applied to design candidate vaccines for other HIV bnAbs or protective Abs against other pathogens.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: WRS is a co-founder and stock holder in Compuvax, Inc. which has programs in non-HIV vaccine design that might benefit indirectly from this research.
Figures

References
-
- Bonsignori M, Hwang KK, Chen X, Tsao CY, Morris L, Gray E, et al. Analysis of a clonal lineage of HIV-1 envelope V2/V3 conformational epitope-specific broadly neutralizing antibodies and their inferred unmutated common ancestors. Journal of virology. 2011;85(19):9998–10009. 10.1128/JVI.05045-11 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
