

Stem and progenitor cells of the mammalian olfactory epithelium: Taking poietic license
- PMID: 27560601
- PMCID: PMC5805156
- DOI: 10.1002/cne.24105
Stem and progenitor cells of the mammalian olfactory epithelium: Taking poietic license
Abstract
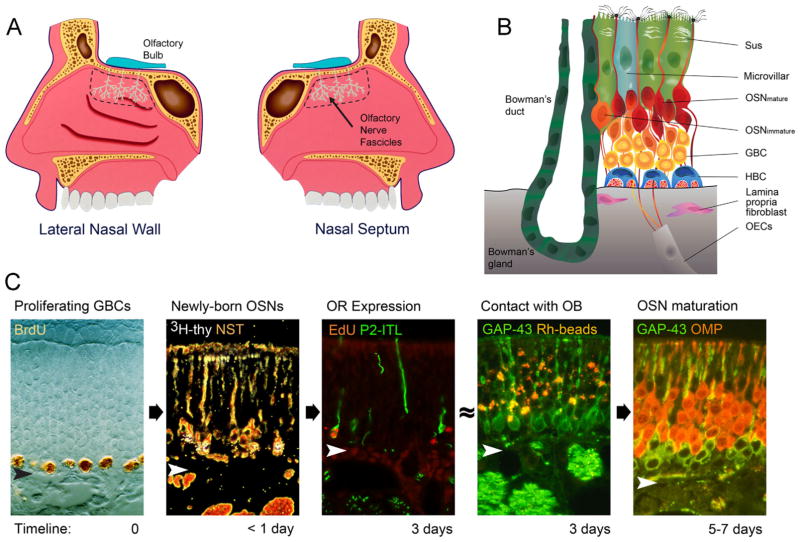
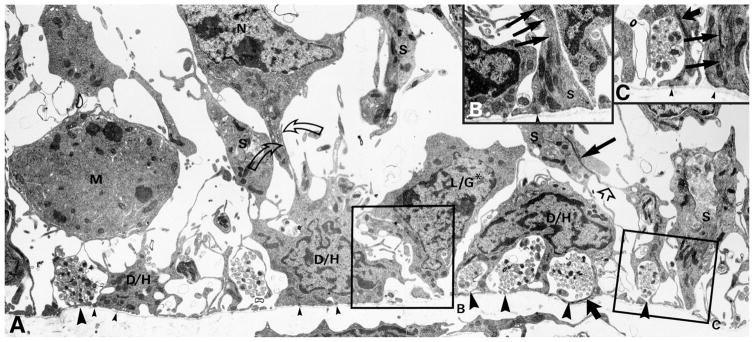
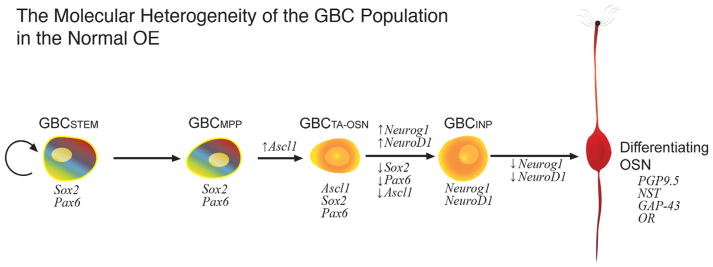
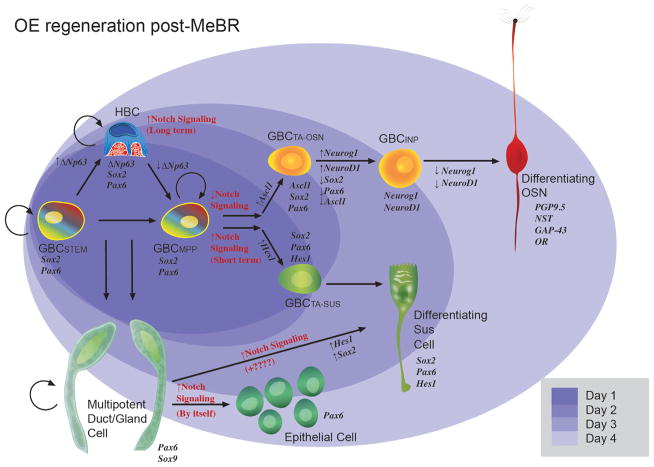
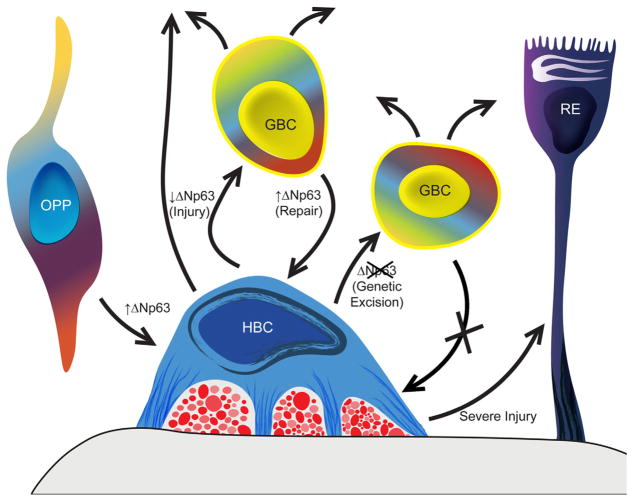
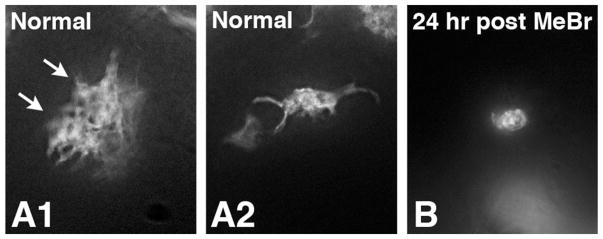
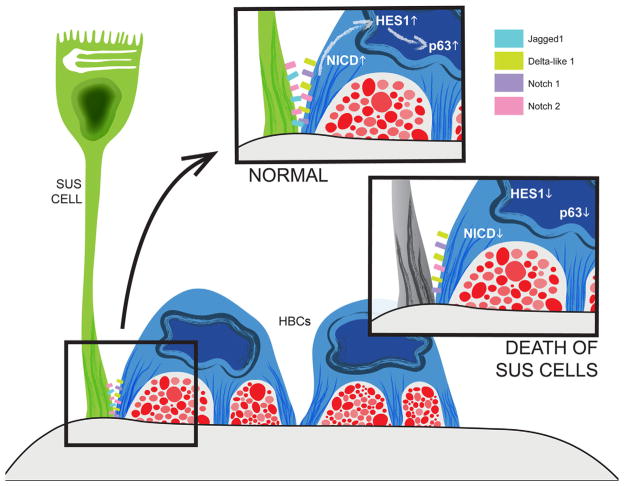
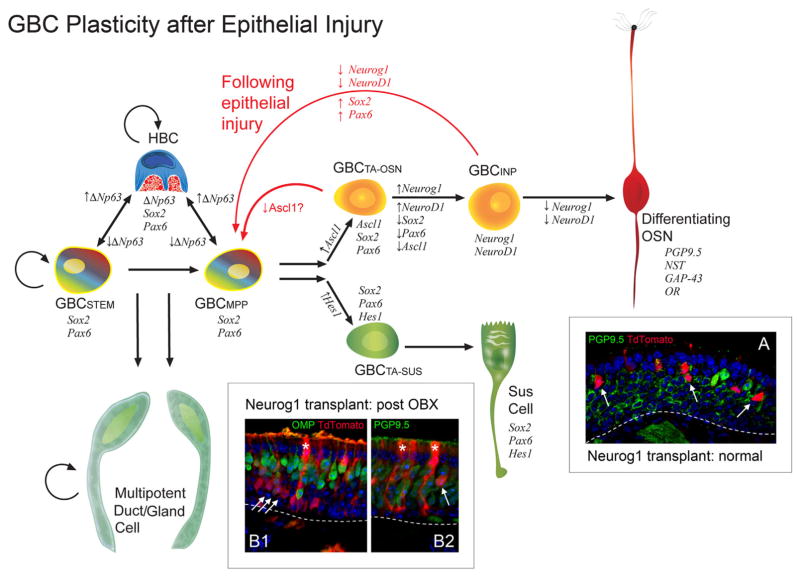
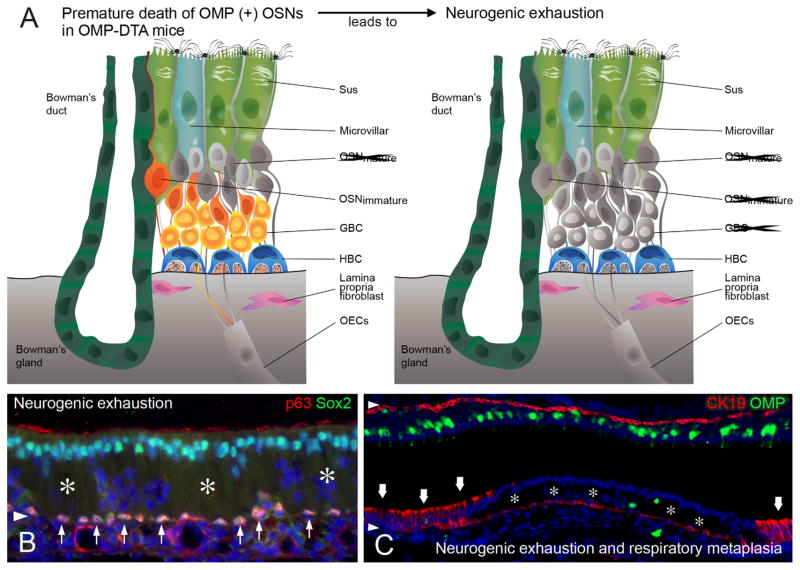
The capacity of the olfactory epithelium (OE) for lifelong neurogenesis and regeneration depends on the persistence of neurocompetent stem cells, which self-renew as well as generating all of the cell types found within the nasal epithelium. This Review focuses on the types of stem and progenitor cells in the epithelium and their regulation. Both horizontal basal cells (HBCs) and some among the population of globose basal cells (GBCs) are stem cells, but the two types plays vastly different roles. The GBC population includes the basal cells that proliferate in the uninjured OE and is heterogeneous with respect to transcription factor expression. From upstream in the hierarchy to downstream, GBCs encompass 1) Sox2+ /Pax6+ stem-like cells that are totipotent and self-renew over the long term, 2) Ascl1+ transit-amplifying progenitors with a limited capacity for expansive proliferation, and 3) Neurog1+ /NeuroD1+ immediate precursor cells that make neurons directly. In contrast, the normally quiescent HBCs are activated to multipotency and proliferate when sustentacular cells are killed, but not when only OSNs die, indicating that HBCs are reserve stem cells that respond to severe epithelial injury. The master regulator of HBC activation is the ΔN isoform of the transcription factor p63; eliminating ΔNp63 unleashes HBC multipotency. Notch signaling, via Jagged1 ligand on Sus cells and Notch1 and Notch2 receptors on HBCs, is likely to play a major role in setting the level of p63 expression. Thus, ΔNp63 becomes a potential therapeutic target for reversing the neurogenic exhaustion characteristic of the aged OE. J. Comp. Neurol. 525:1034-1054, 2017. © 2016 Wiley Periodicals, Inc.
Keywords: active stem cell; aging; dedifferentiation; globose basal cell; horizontal basal cell; neurogenesis; p63; reserve stem cell.
© 2016 Wiley Periodicals, Inc.
Conflict of interest statement
The authors have no conflicts of interest.
Figures
References
-
- Andres KH. The fine structure of the olfactory region of macrosmatic animals. Z Zellforsch Mikrosk Anat. 1966;69:140–154. - PubMed
-
- Asan E, Drenckhahn D. Immunocytochemical characterization of two types of microvillar cells in rodent olfactory epithelium. Histochem Cell Biol. 2005;123:157–168. - PubMed
-
- Becker AJ, McCulloch EA, Till JE. Cytological demonstration of the clonal nature of spleen colonies derived from transplanted mouse marrow cells. Nature. 1963;197:452–454. - PubMed
-
- Bergman U, Ostergren A, Gustafson AL, Brittebo B. Differential effects of olfactory toxicants on olfactory regeneration. Arch Toxicol. 2002;76:104–112. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
