

Cell non-autonomous regulation of hepatic IGF-1 and neonatal growth by Kinase Suppressor of Ras 2 (KSR2)
- PMID: 27561547
- PMCID: PMC4999994
- DOI: 10.1038/srep32093
Cell non-autonomous regulation of hepatic IGF-1 and neonatal growth by Kinase Suppressor of Ras 2 (KSR2)
Abstract
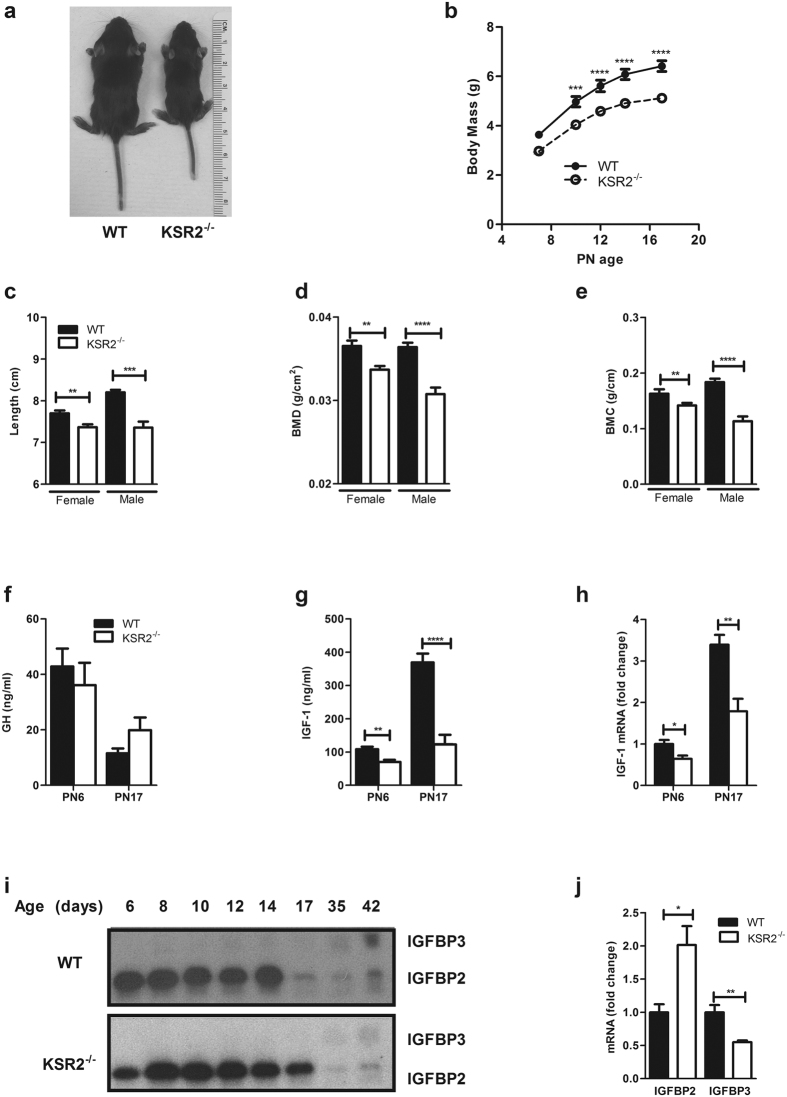
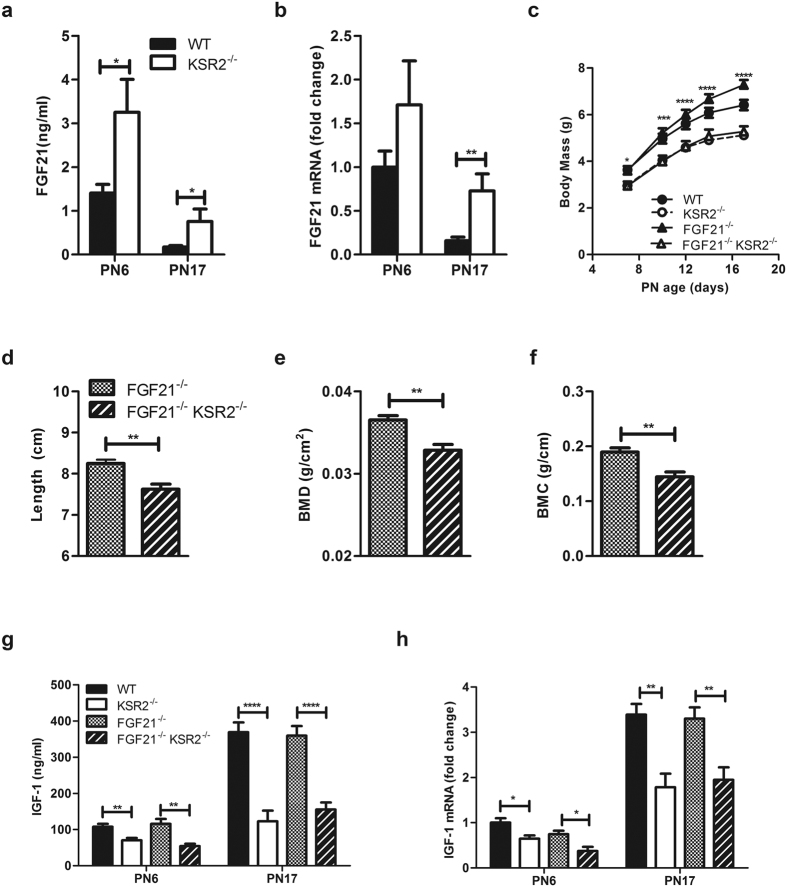
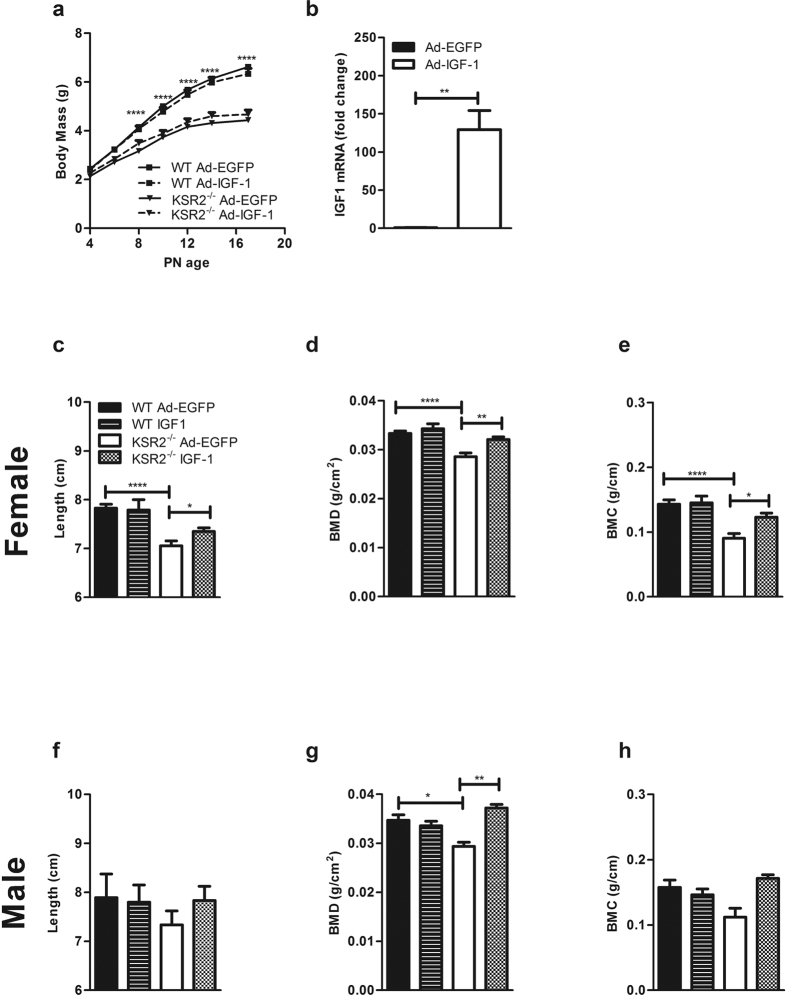
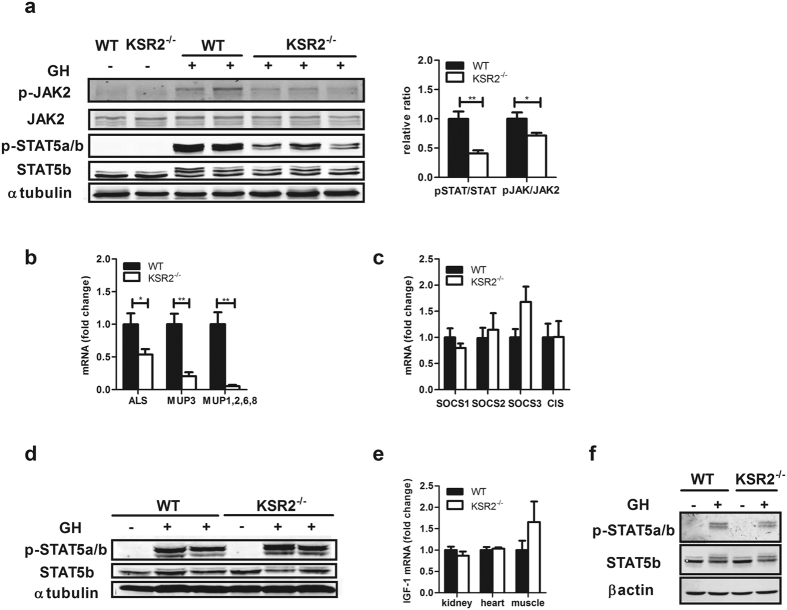
Individuals with poor postnatal growth are at risk for cardiovascular and metabolic problems as adults. Here we show that disruption of the molecular scaffold Kinase Suppressor of Ras 2 (KSR2) causes selective inhibition of hepatic GH signaling in neonatal mice with impaired expression of IGF-1 and IGFBP3. ksr2(-/-) mice are normal size at birth but show a marked increase in FGF21 accompanied by reduced body mass, shortened body length, and reduced bone mineral density (BMD) and content (BMC) first evident during postnatal development. However, disrupting FGF21 in ksr2(-/-) mice does not normalize mass, length, or bone density and content in fgf21(-/-)ksr2(-/-) mice. Body length, BMC and BMD, but not body mass, are rescued by infection of two-day-old ksr2(-/-) mice with a recombinant adenovirus encoding human IGF-1. Relative to wild-type mice, GH injections reveal a significant reduction in JAK2 and STAT5 phosphorylation in liver, but not in skeletal muscle, of ksr2(-/-) mice. However, primary hepatocytes isolated from ksr2(-/-) mice show no reduction in GH-stimulated STAT5 phosphorylation. These data indicate that KSR2 functions in a cell non-autonomous fashion to regulate GH-stimulated IGF-1 expression in the liver of neonatal mice, which plays a key role in the development of body length.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
