

Na+ coordination at the Na2 site of the Na+/I- symporter
- PMID: 27562170
- PMCID: PMC5027462
- DOI: 10.1073/pnas.1607231113
Na+ coordination at the Na2 site of the Na+/I- symporter
Abstract
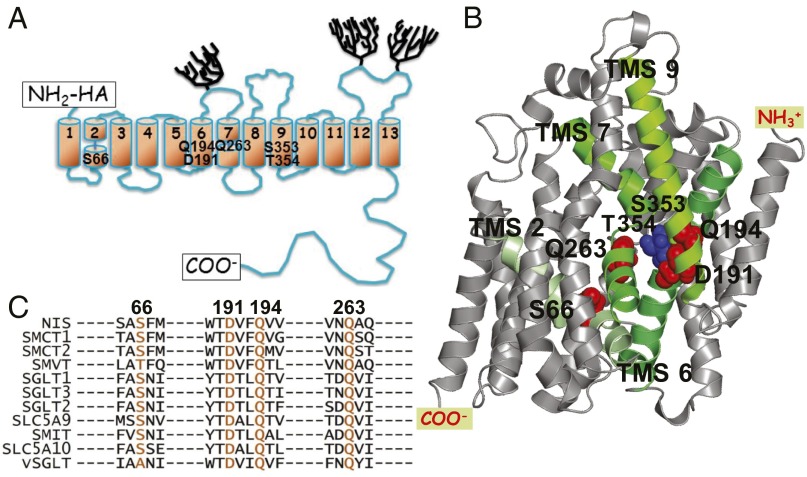
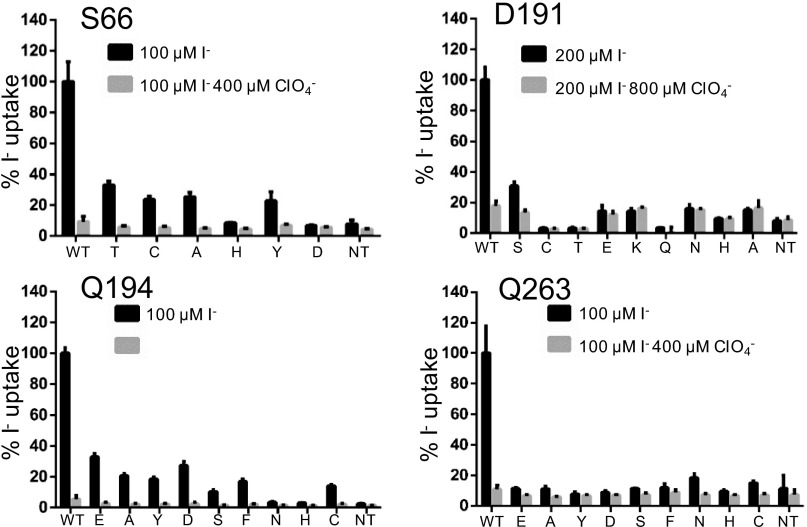
The sodium/iodide symporter (NIS) mediates active I(-) transport in the thyroid-the first step in thyroid hormone biosynthesis-with a 2 Na(+): 1 I(-) stoichiometry. The two Na(+) binding sites (Na1 and Na2) and the I(-) binding site interact allosterically: when Na(+) binds to a Na(+) site, the affinity of NIS for the other Na(+) and for I(-) increases significantly. In all Na(+)-dependent transporters with the same fold as NIS, the side chains of two residues, S353 and T354 (NIS numbering), were identified as the Na(+) ligands at Na2. To understand the cooperativity between the substrates, we investigated the coordination at the Na2 site. We determined that four other residues-S66, D191, Q194, and Q263-are also involved in Na(+) coordination at this site. Experiments in whole cells demonstrated that these four residues participate in transport by NIS: mutations at these positions result in proteins that, although expressed at the plasma membrane, transport little or no I(-) These residues are conserved throughout the entire SLC5 family, to which NIS belongs, suggesting that they serve a similar function in the other transporters. Our findings also suggest that the increase in affinity that each site displays when an ion binds to another site may result from changes in the dynamics of the transporter. These mechanistic insights deepen our understanding not only of NIS but also of other transporters, including many that, like NIS, are of great medical relevance.
Keywords: NIS; Na+ binding site; Na+-driven cotransporters; Na+/I− symporter; protein dynamics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures











References
-
- Carrasco N. Iodide transport in the thyroid gland. Biochim Biophys Acta. 1993;1154(1):65–82. - PubMed
-
- Dai G, Levy O, Carrasco N. Cloning and characterization of the thyroid iodide transporter. Nature. 1996;379(6564):458–460. - PubMed
-
- De La Vieja A, Dohan O, Levy O, Carrasco N. Molecular analysis of the sodium/iodide symporter: Impact on thyroid and extrathyroid pathophysiology. Physiol Rev. 2000;80(3):1083–1105. - PubMed
-
- Eskandari S, et al. Thyroid Na+/I− symporter. Mechanism, stoichiometry, and specificity. J Biol Chem. 1997;272(43):27230–27238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
