

Evolution and Molecular Control of Hybrid Incompatibility in Plants
- PMID: 27563306
- PMCID: PMC4980391
- DOI: 10.3389/fpls.2016.01208
Evolution and Molecular Control of Hybrid Incompatibility in Plants
Abstract
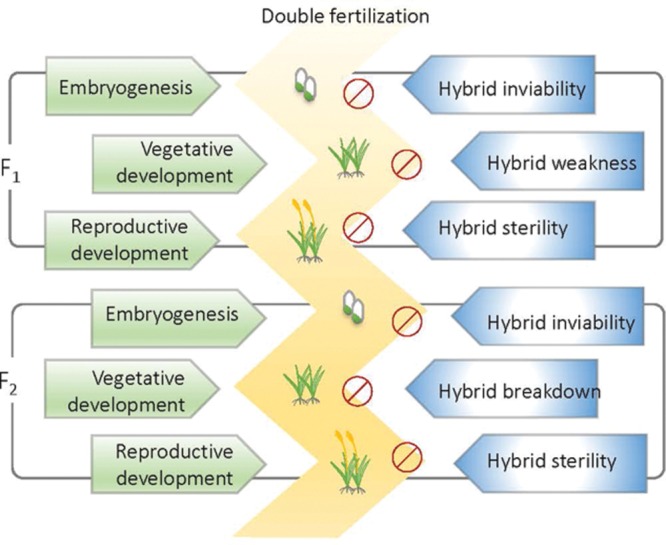
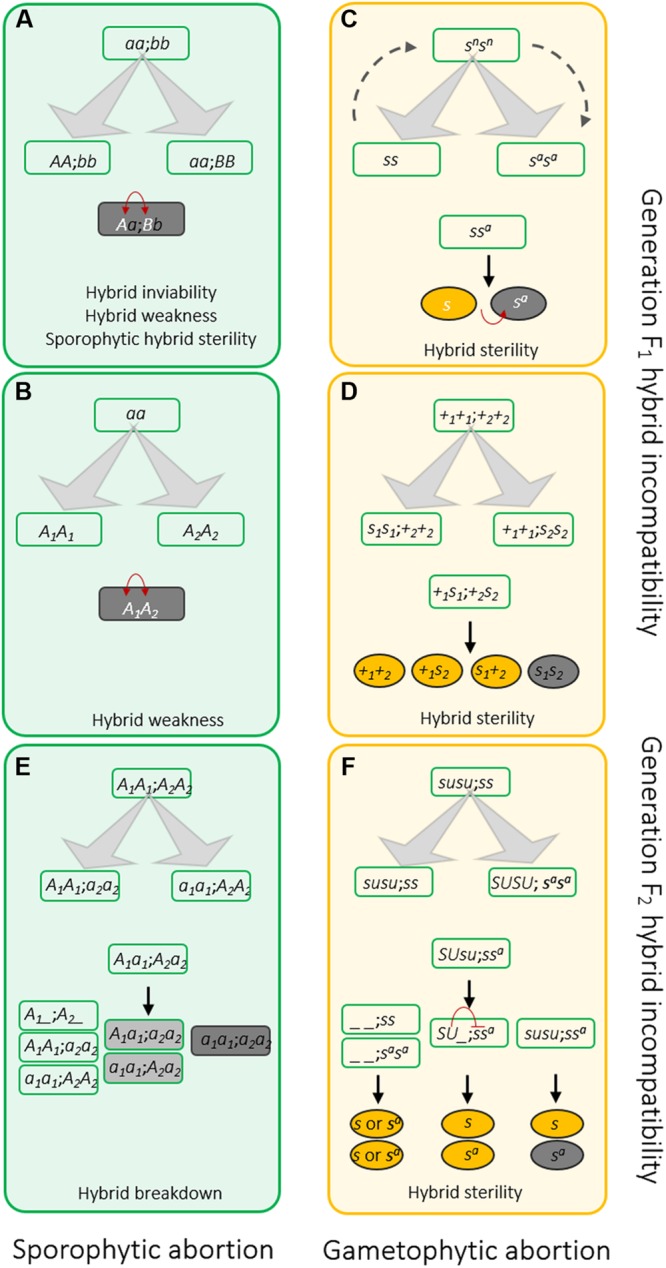
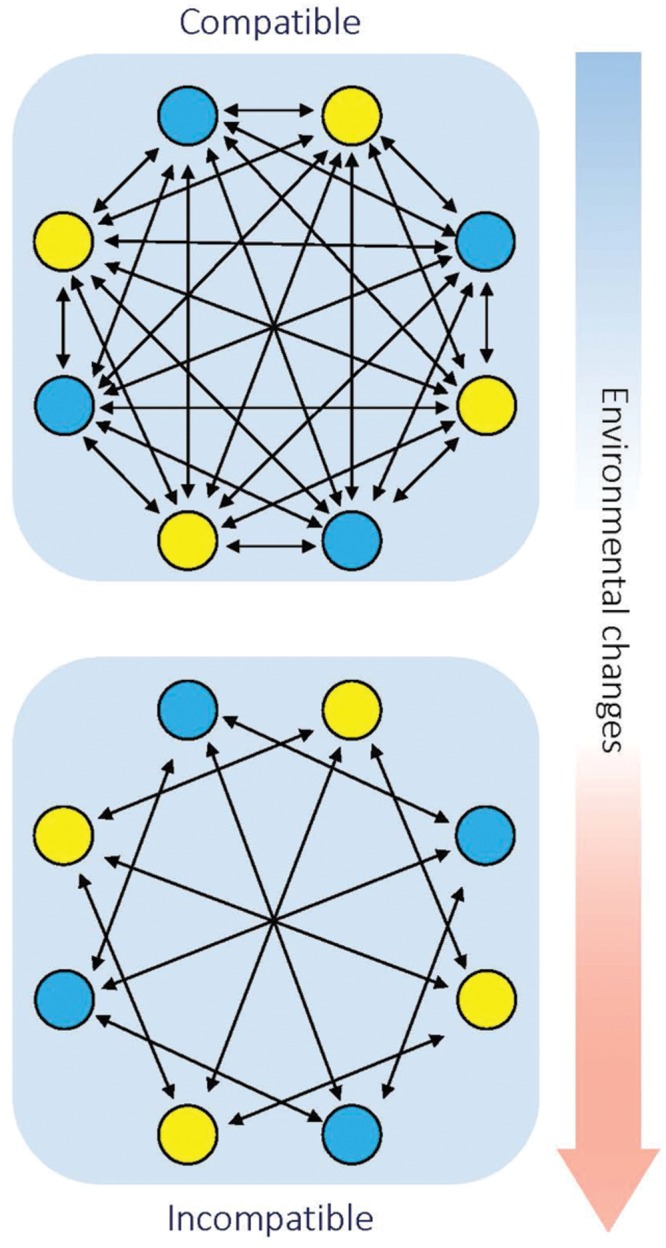
Postzygotic reproductive isolation (RI) plays an important role in speciation. According to the stage at which it functions and the symptoms it displays, postzygotic RI can be called hybrid inviability, hybrid weakness or necrosis, hybrid sterility, or hybrid breakdown. In this review, we summarized new findings about hybrid incompatibilities in plants, most of which are from studies on Arabidopsis and rice. Recent progress suggests that hybrid incompatibility is a by-product of co-evolution either with "parasitic" selfish elements in the genome or with invasive microbes in the natural environment. We discuss the environmental influences on the expression of hybrid incompatibility and the possible effects of environment-dependent hybrid incompatibility on sympatric speciation. We also discuss the role of domestication on the evolution of hybrid incompatibilities.
Keywords: Bateson-Dobzhansky–Muller model; Hybrid incompatibility; Speciation genes; evolutionary force; evolutionary genetics; postzygotic reproductive barriers.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources