

Seipin is required for converting nascent to mature lipid droplets
- PMID: 27564575
- PMCID: PMC5035145
- DOI: 10.7554/eLife.16582
Seipin is required for converting nascent to mature lipid droplets
Abstract
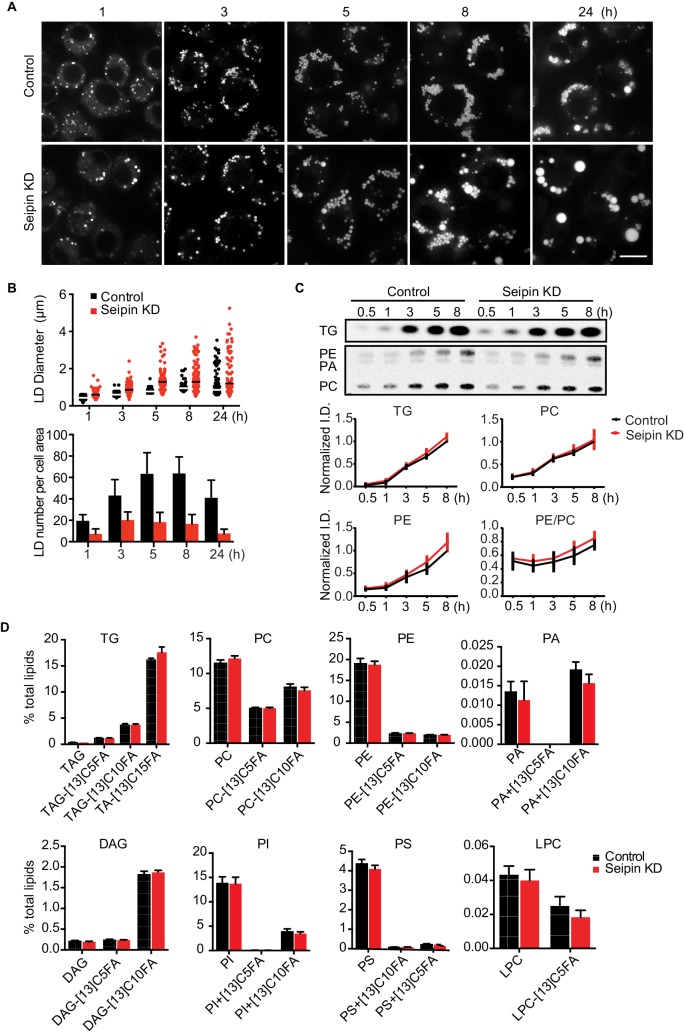
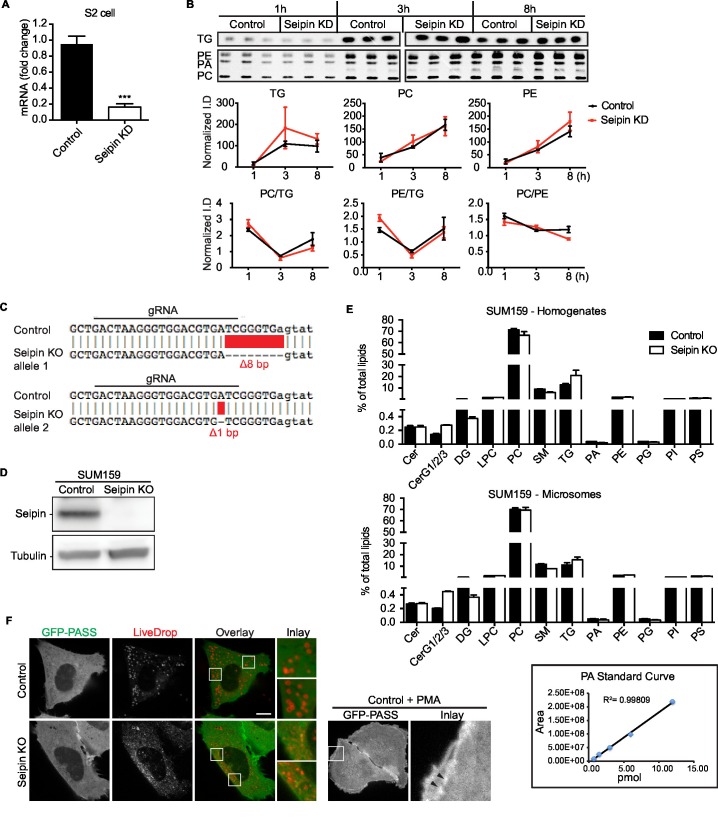
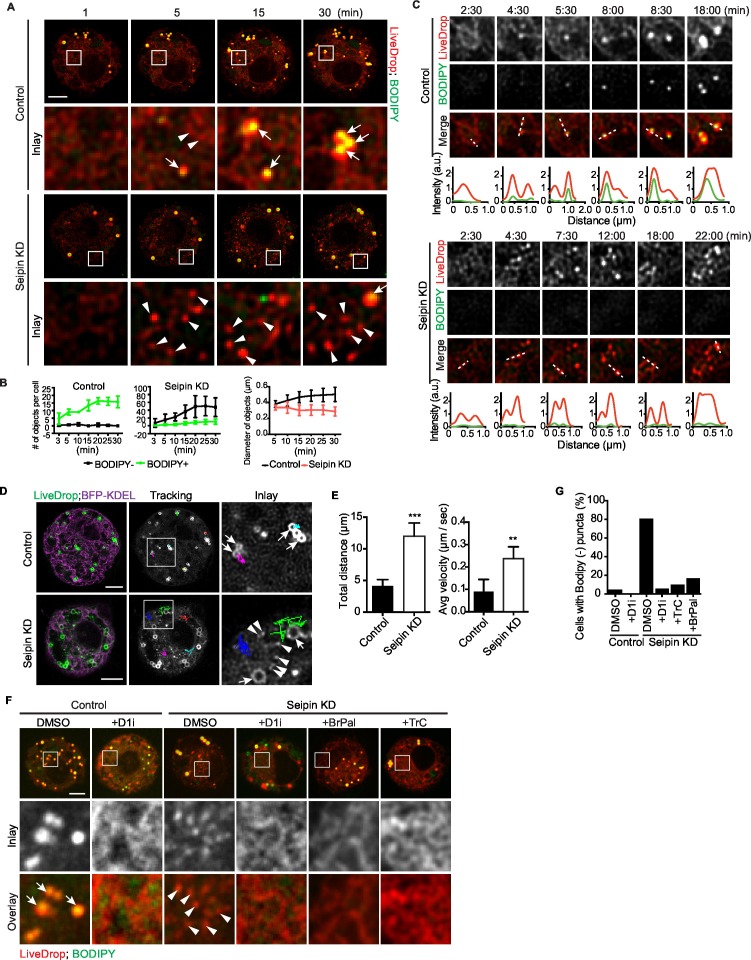
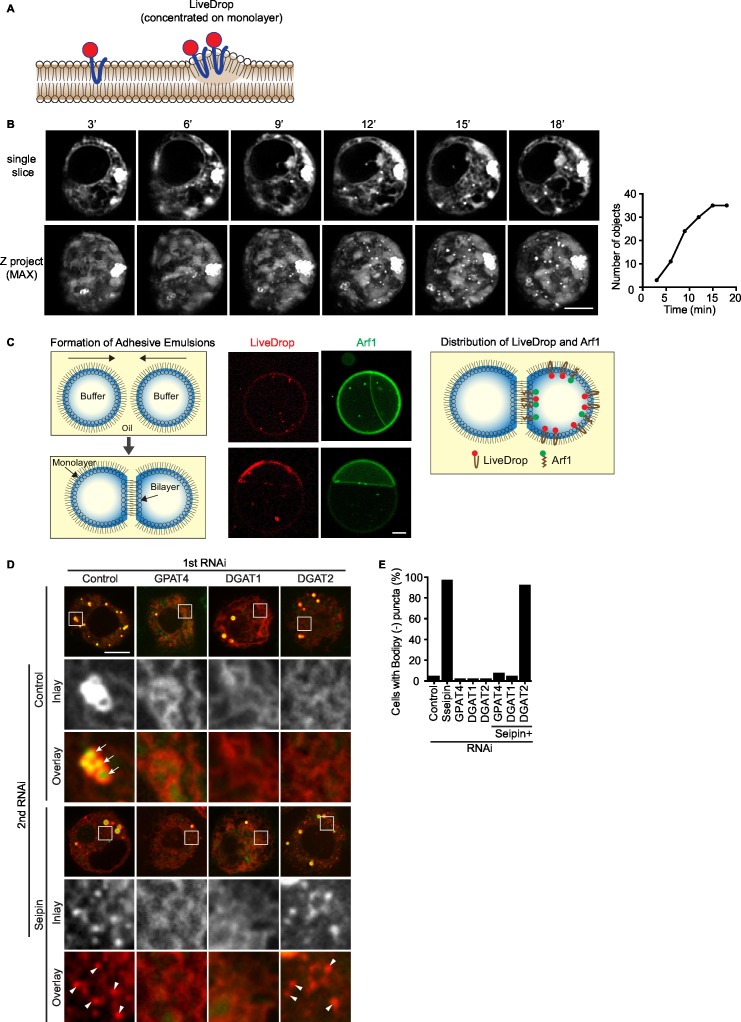
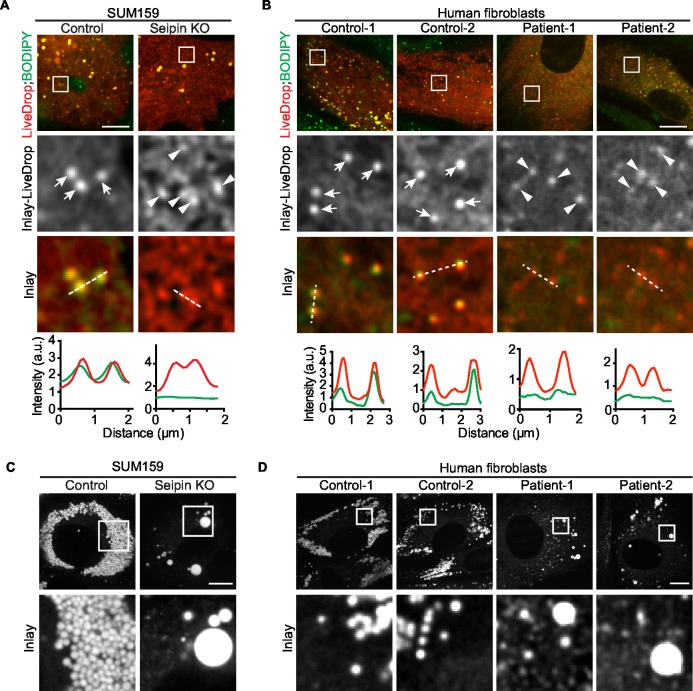
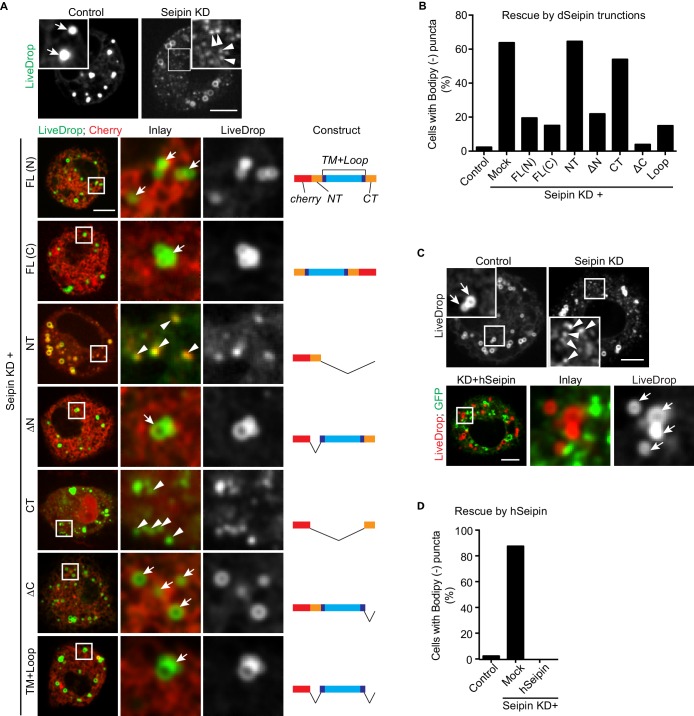
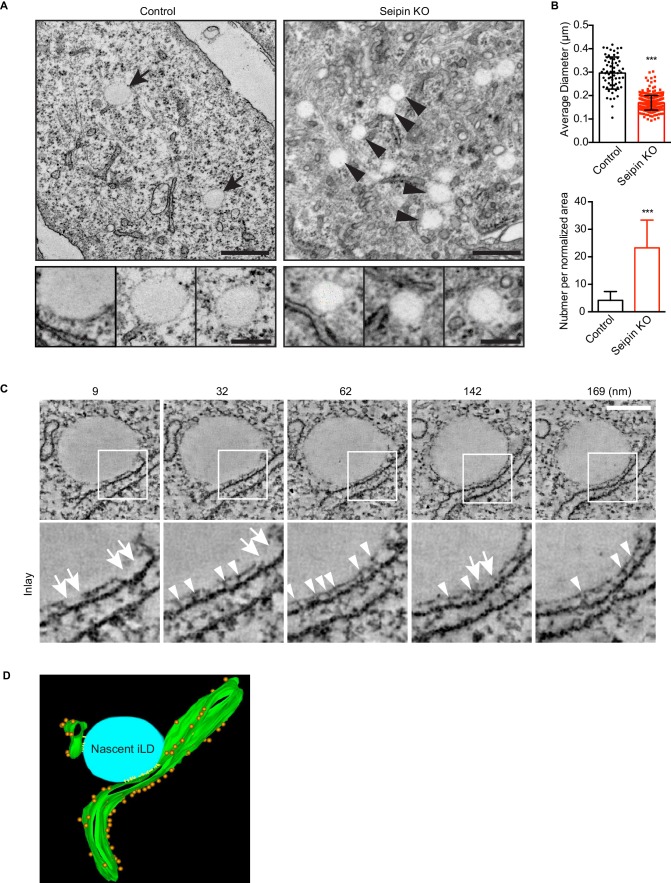
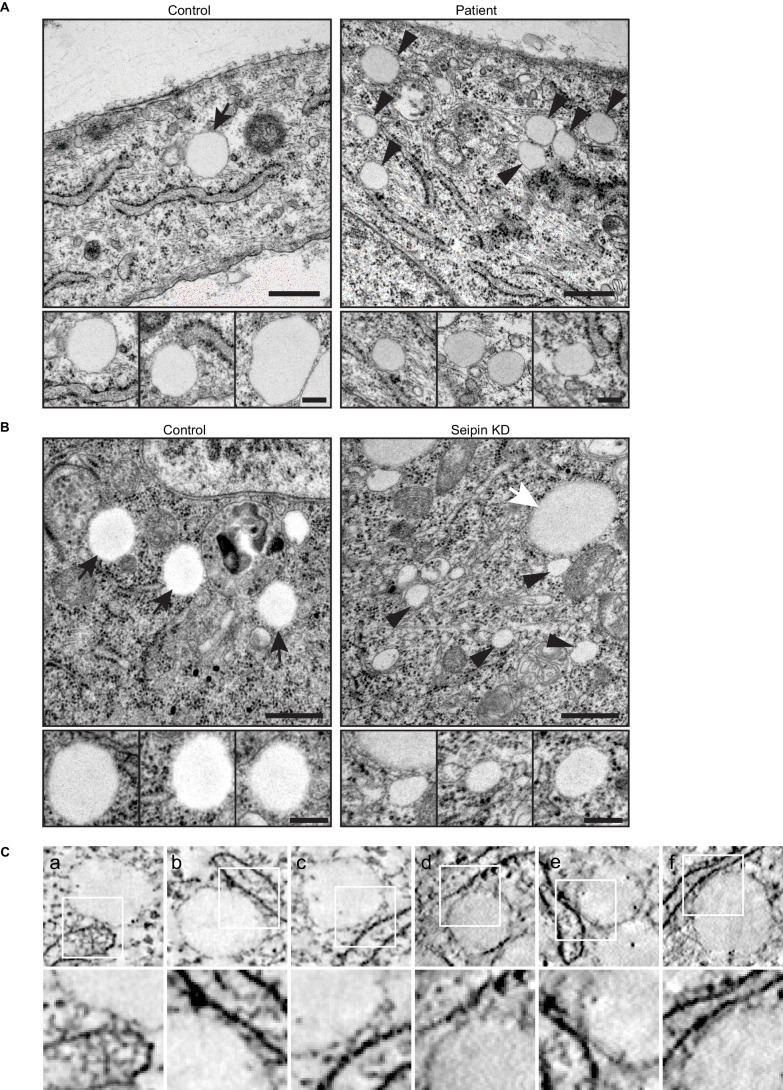
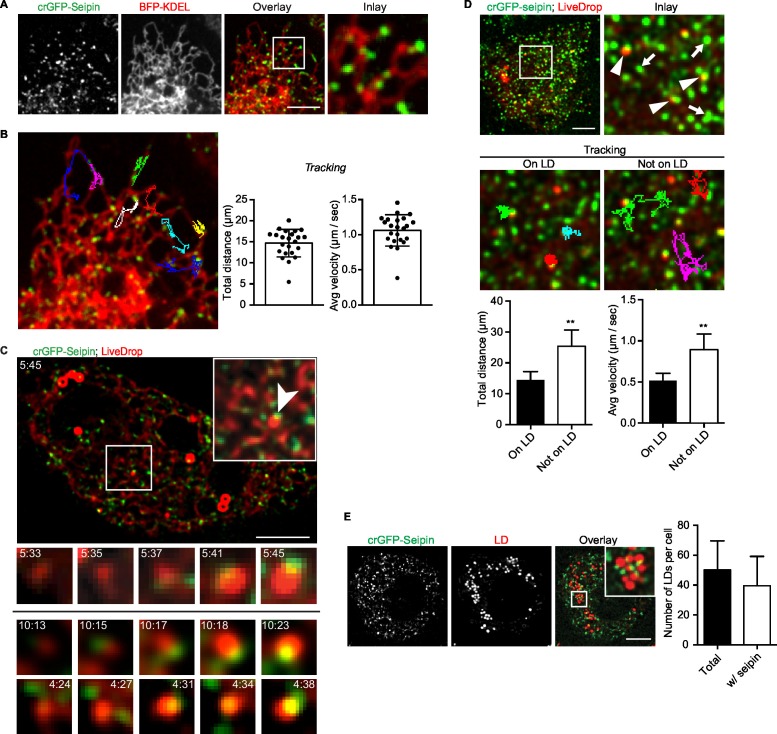
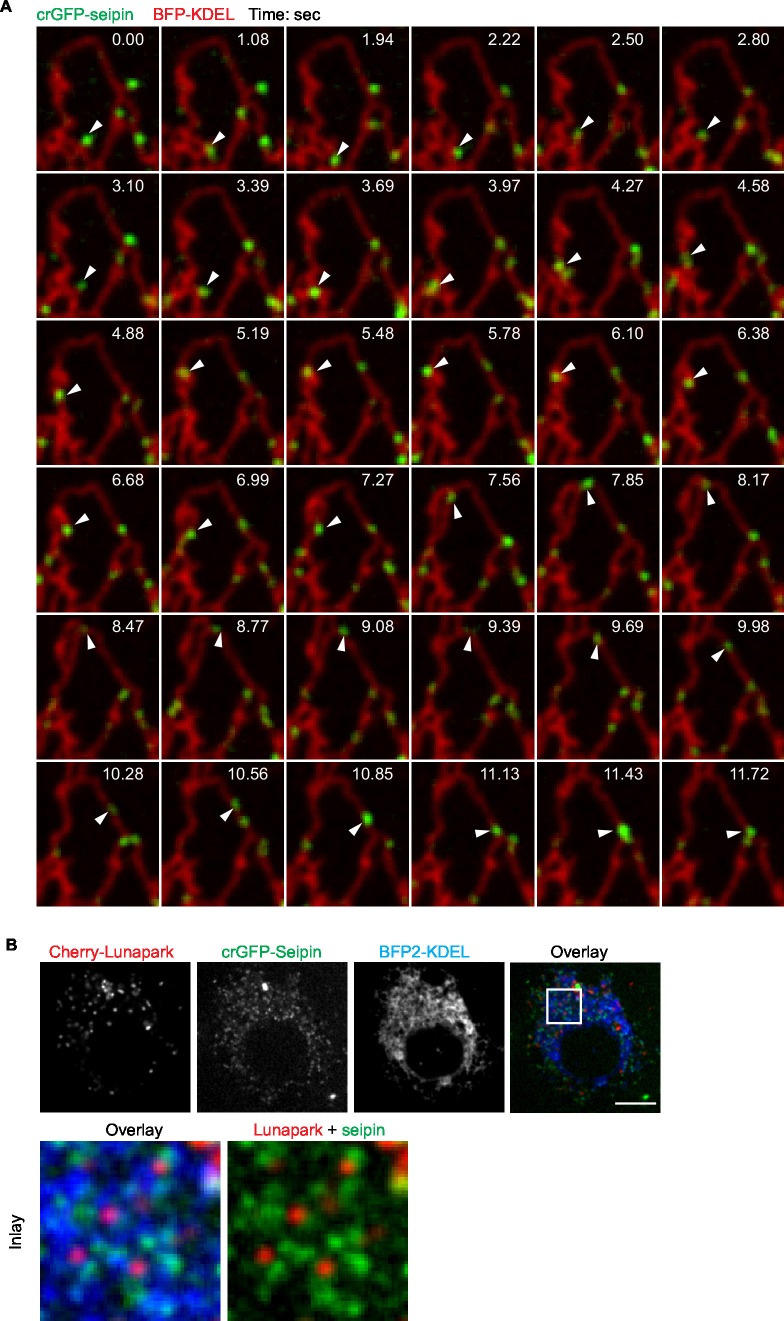
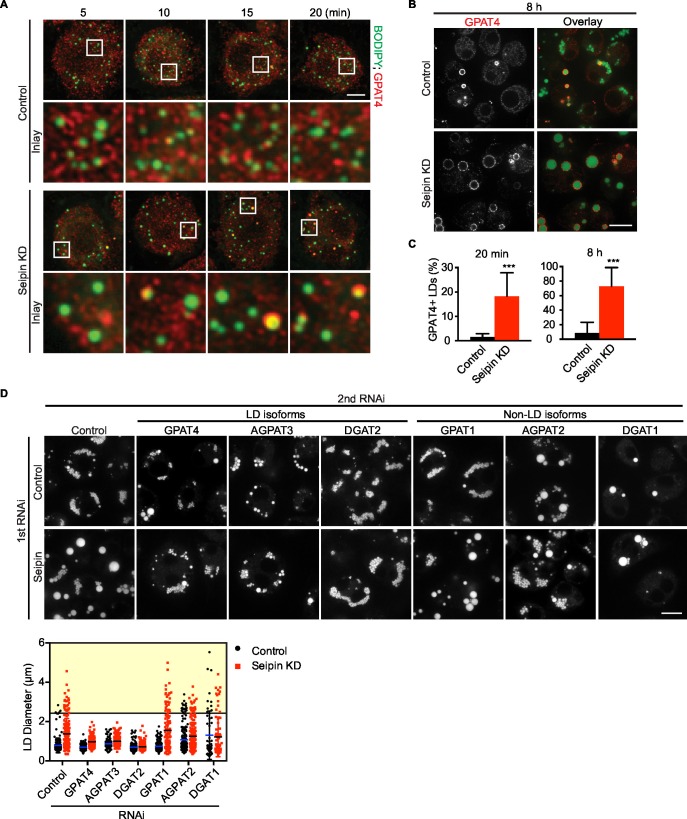
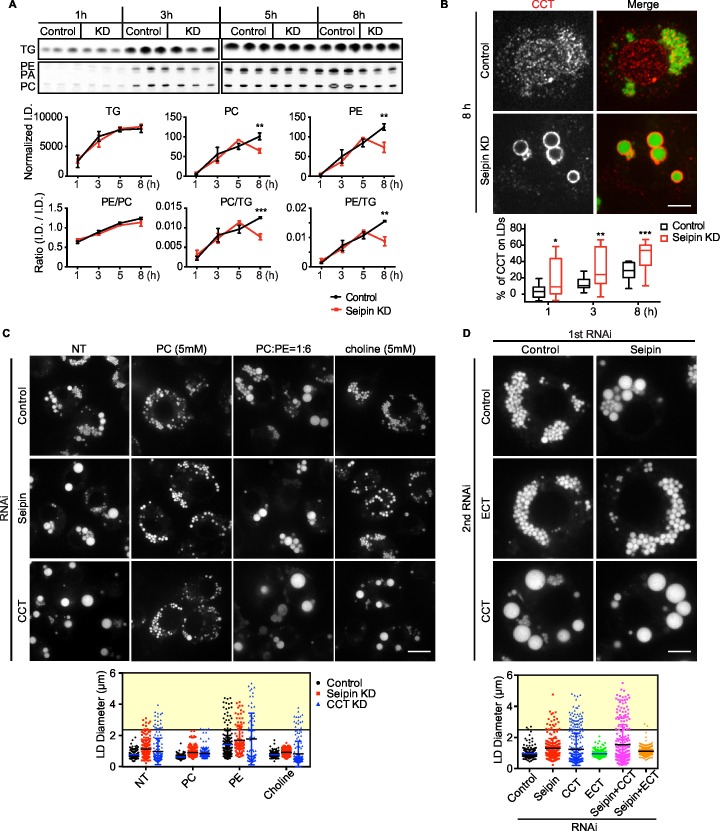
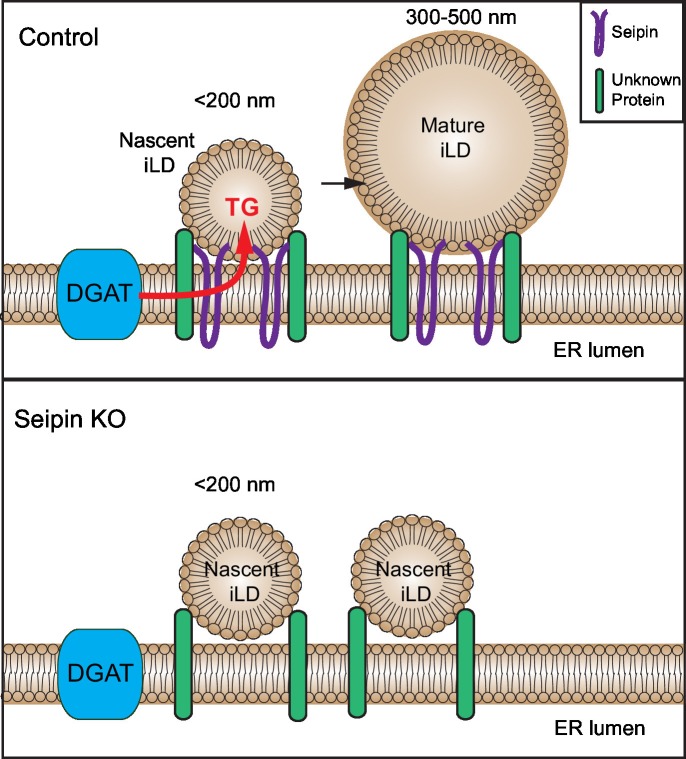
How proteins control the biogenesis of cellular lipid droplets (LDs) is poorly understood. Using Drosophila and human cells, we show here that seipin, an ER protein implicated in LD biology, mediates a discrete step in LD formation-the conversion of small, nascent LDs to larger, mature LDs. Seipin forms discrete and dynamic foci in the ER that interact with nascent LDs to enable their growth. In the absence of seipin, numerous small, nascent LDs accumulate near the ER and most often fail to grow. Those that do grow prematurely acquire lipid synthesis enzymes and undergo expansion, eventually leading to the giant LDs characteristic of seipin deficiency. Our studies identify a discrete step of LD formation, namely the conversion of nascent LDs to mature LDs, and define a molecular role for seipin in this process, most likely by acting at ER-LD contact sites to enable lipid transfer to nascent LDs.
Keywords: D. melanogaster; LiveDrop; cell biology; endoplasmic reticulum; human; lipid droplet; lipid metabolism; organelle biogenesis; seipin.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures


References
-
- Agarwal AK, Simha V, Oral EA, Moran SA, Gorden P, O'Rahilly S, Zaidi Z, Gurakan F, Arslanian SA, Klar A, Ricker A, White NH, Bindl L, Herbst K, Kennel K, Patel SB, Al-Gazali L, Garg A. Phenotypic and genetic heterogeneity in congenital generalized lipodystrophy. The Journal of Clinical Endocrinology & Metabolism. 2003;88:4840–4847. doi: 10.1210/jc.2003-030855. - DOI - PubMed
-
- Almeida R, Pauling JK, Sokol E, Hannibal-Bach HK, Ejsing CS. Comprehensive lipidome analysis by shotgun lipidomics on a hybrid quadrupole-orbitrap-linear ion trap mass spectrometer. Journal of the American Society for Mass Spectrometry. 2015;26:133–148. doi: 10.1007/s13361-014-1013-x. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
