

Cell-Type-Specific Alternative Splicing Governs Cell Fate in the Developing Cerebral Cortex
- PMID: 27565344
- PMCID: PMC5248659
- DOI: 10.1016/j.cell.2016.07.025
Cell-Type-Specific Alternative Splicing Governs Cell Fate in the Developing Cerebral Cortex
Abstract
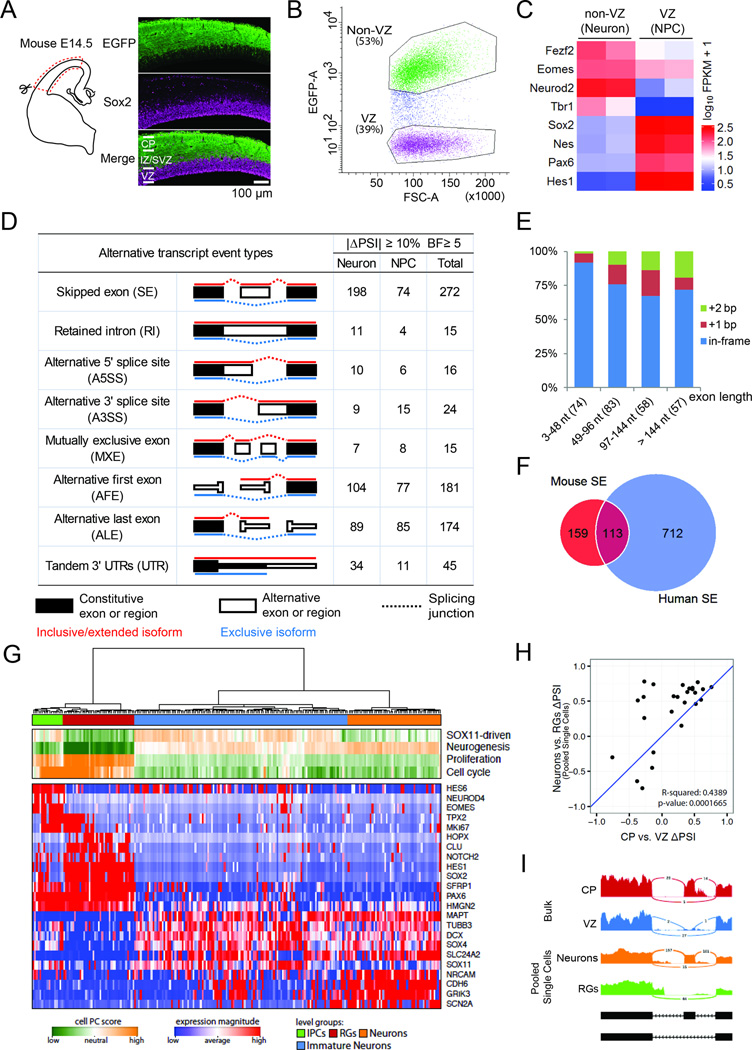
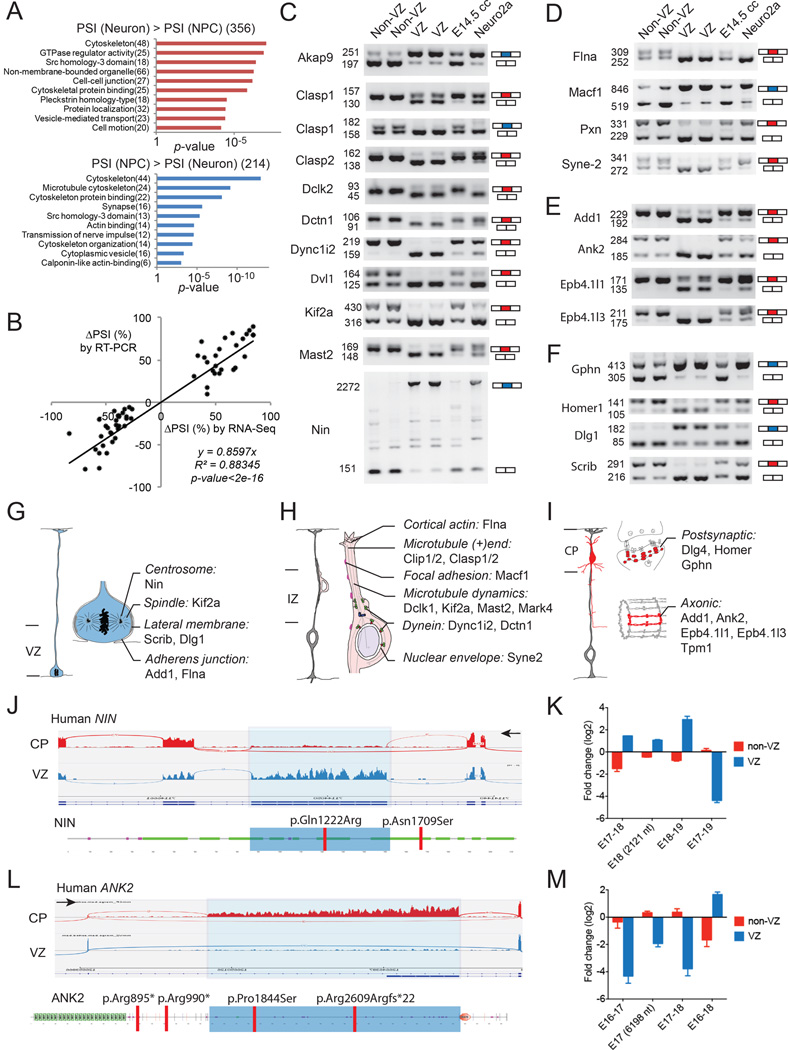
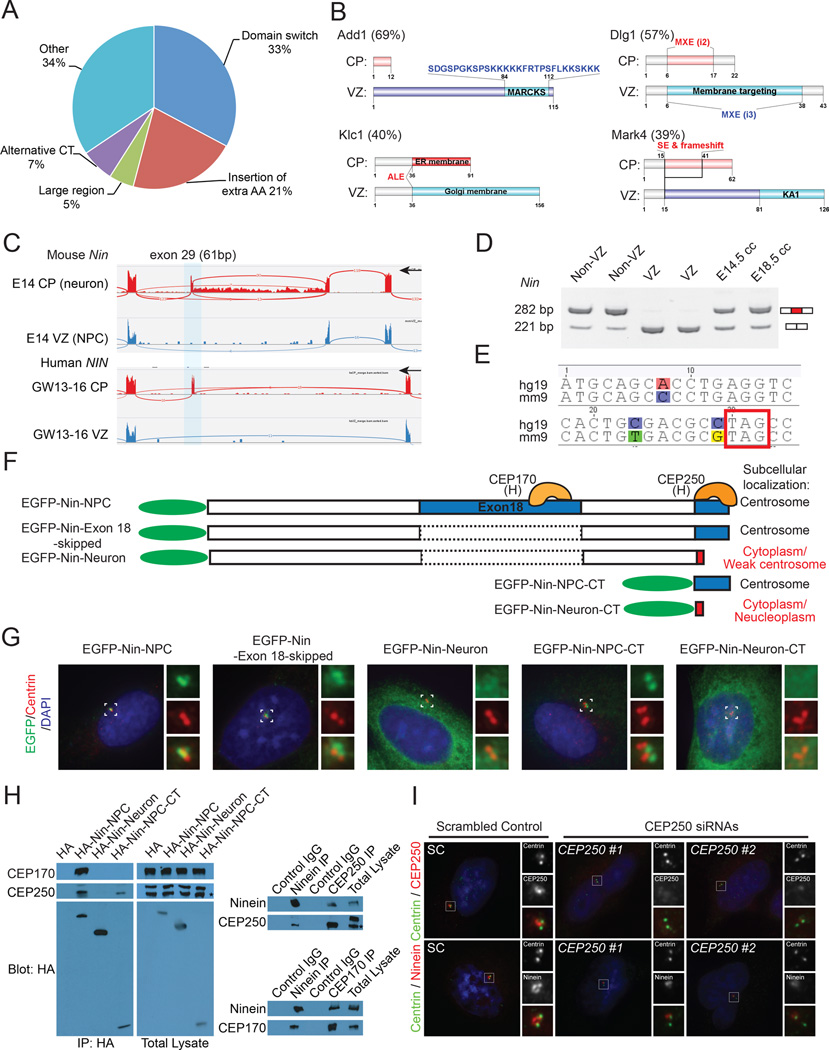
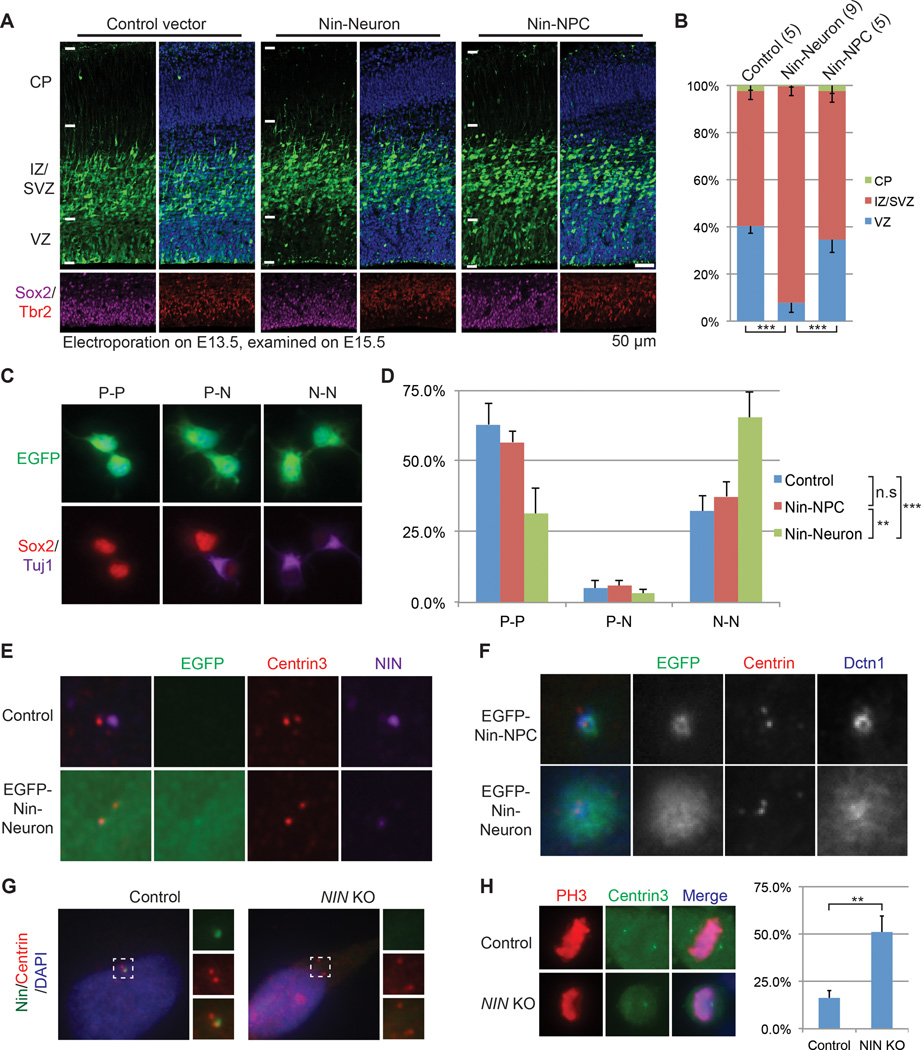
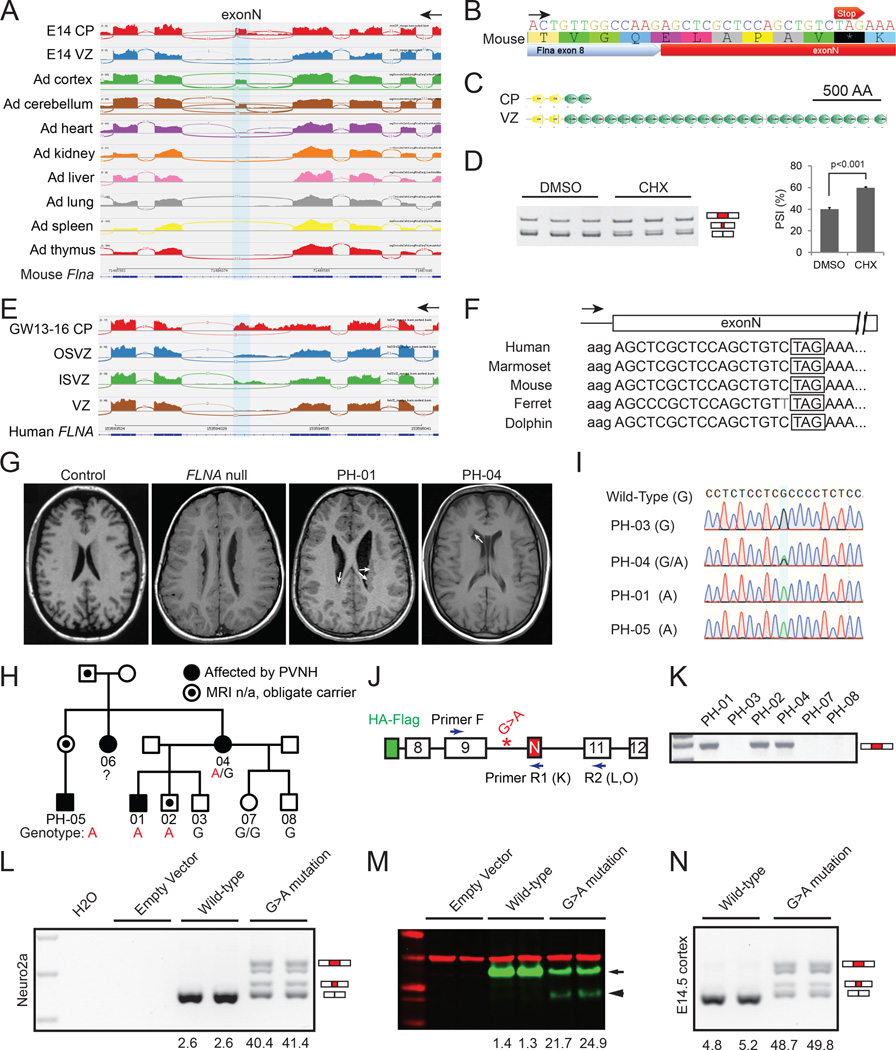
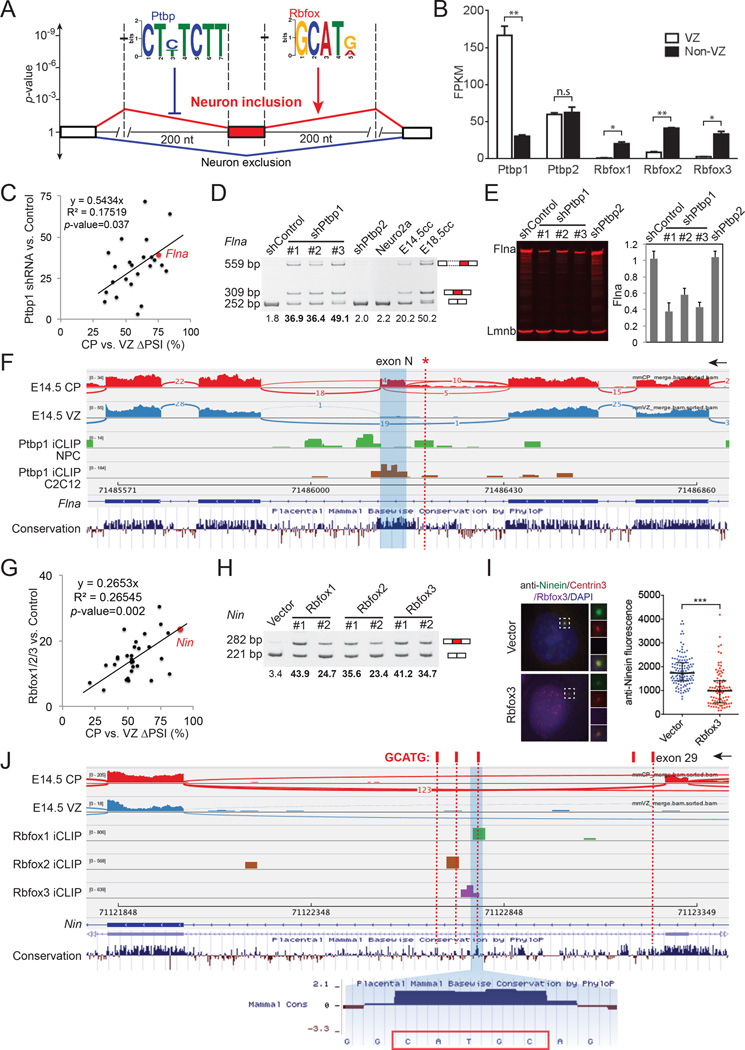
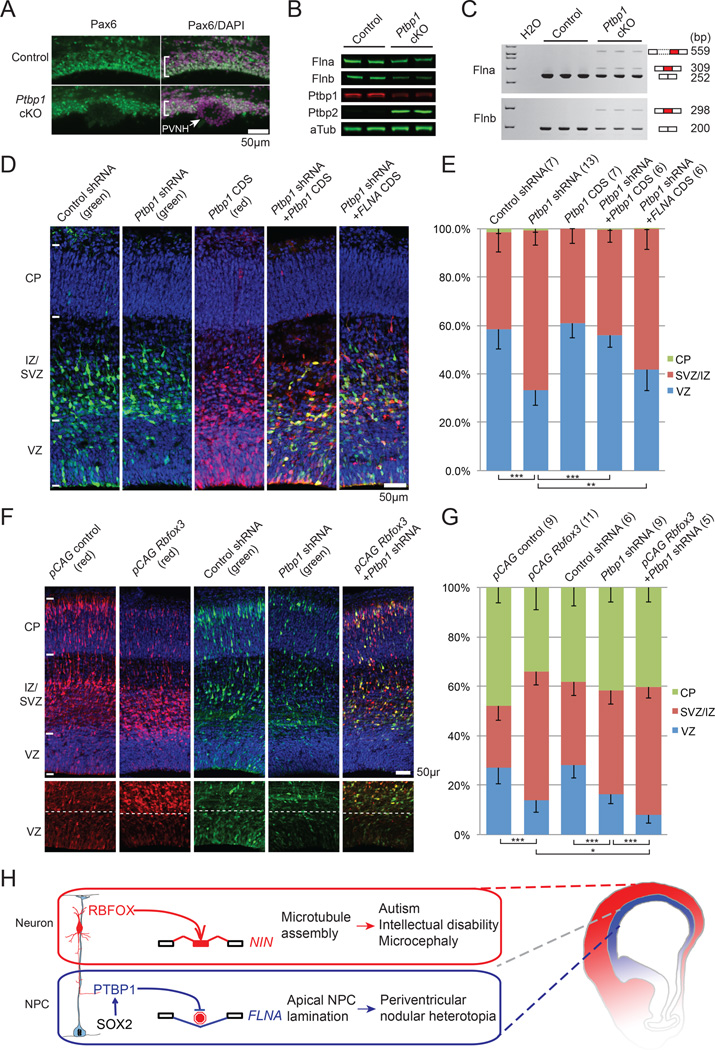
Alternative splicing is prevalent in the mammalian brain. To interrogate the functional role of alternative splicing in neural development, we analyzed purified neural progenitor cells (NPCs) and neurons from developing cerebral cortices, revealing hundreds of differentially spliced exons that preferentially alter key protein domains-especially in cytoskeletal proteins-and can harbor disease-causing mutations. We show that Ptbp1 and Rbfox proteins antagonistically govern the NPC-to-neuron transition by regulating neuron-specific exons. Whereas Ptbp1 maintains apical progenitors partly through suppressing a poison exon of Flna in NPCs, Rbfox proteins promote neuronal differentiation by switching Ninein from a centrosomal splice form in NPCs to a non-centrosomal isoform in neurons. We further uncover an intronic human mutation within a PTBP1-binding site that disrupts normal skipping of the FLNA poison exon in NPCs and causes a brain-specific malformation. Our study indicates that dynamic control of alternative splicing governs cell fate in cerebral cortical development.
Keywords: Ninein; Ptbp1; Rbfox; filamin A; microcephaly; mother centriole; periventricular nodular heterotopia.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Ayoub AE, Oh S, Xie Y, Leng J, Cotney J, Dominguez MH, Noonan JP, Rakic P. Transcriptional programs in transient embryonic zones of the cerebral cortex defined by high-resolution mRNA sequencing. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:14950–14955. - PMC - PubMed
-
- Baird DH, Myers KA, Mogensen M, Moss D, Baas PW. Distribution of the microtubule-related protein ninein in developing neurons. Neuropharmacology. 2004;47:677–683. - PubMed
-
- Barbosa-Morais NL, Irimia M, Pan Q, Xiong HY, Gueroussov S, Lee LJ, Slobodeniuc V, Kutter C, Watt S, Colak R, et al. The evolutionary landscape of alternative splicing in vertebrate species. Science. 2012;338:1587–1593. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
