

Two Distinct Types of E3 Ligases Work in Unison to Regulate Substrate Ubiquitylation
- PMID: 27565346
- PMCID: PMC5091668
- DOI: 10.1016/j.cell.2016.07.027
Two Distinct Types of E3 Ligases Work in Unison to Regulate Substrate Ubiquitylation
Abstract
Hundreds of human cullin-RING E3 ligases (CRLs) modify thousands of proteins with ubiquitin (UB) to achieve vast regulation. Current dogma posits that CRLs first catalyze UB transfer from an E2 to their client substrates and subsequent polyubiquitylation from various linkage-specific E2s. We report an alternative E3-E3 tagging cascade: many cellular NEDD8-modified CRLs associate with a mechanistically distinct thioester-forming RBR-type E3, ARIH1, and rely on ARIH1 to directly add the first UB and, in some cases, multiple additional individual monoubiquitin modifications onto CRL client substrates. Our data define ARIH1 as a component of the human CRL system, demonstrate that ARIH1 can efficiently and specifically mediate monoubiquitylation of several CRL substrates, and establish principles for how two distinctive E3s can reciprocally control each other for simultaneous and joint regulation of substrate ubiquitylation. These studies have broad implications for CRL-dependent proteostasis and mechanisms of E3-mediated UB ligation.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
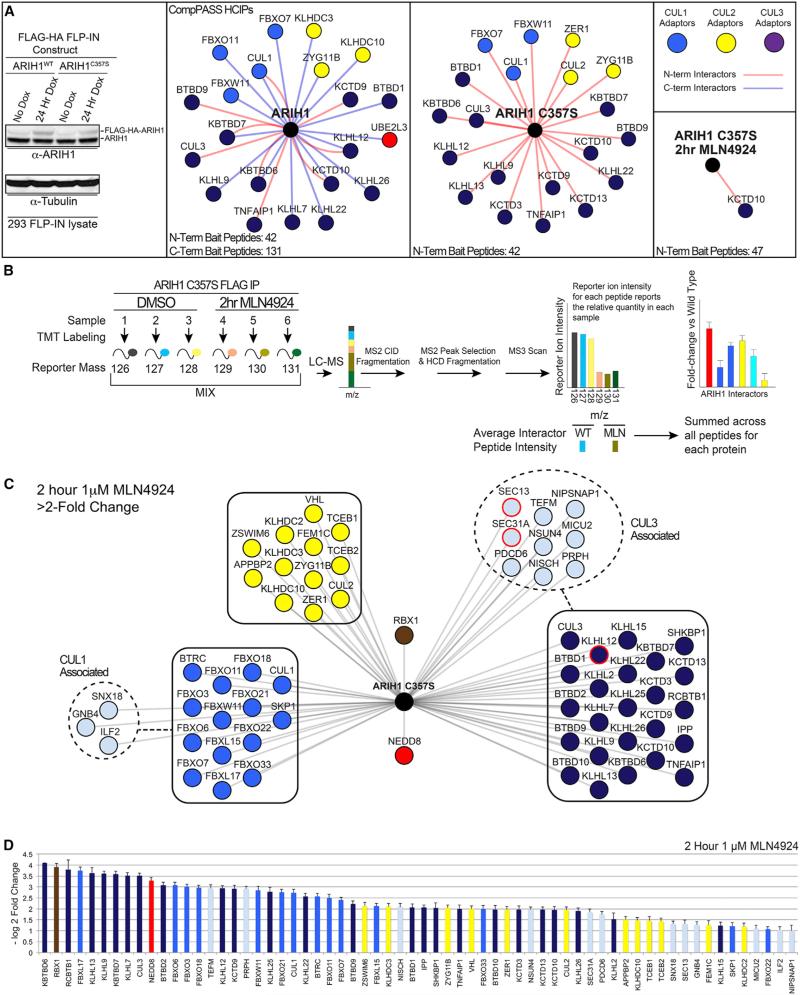

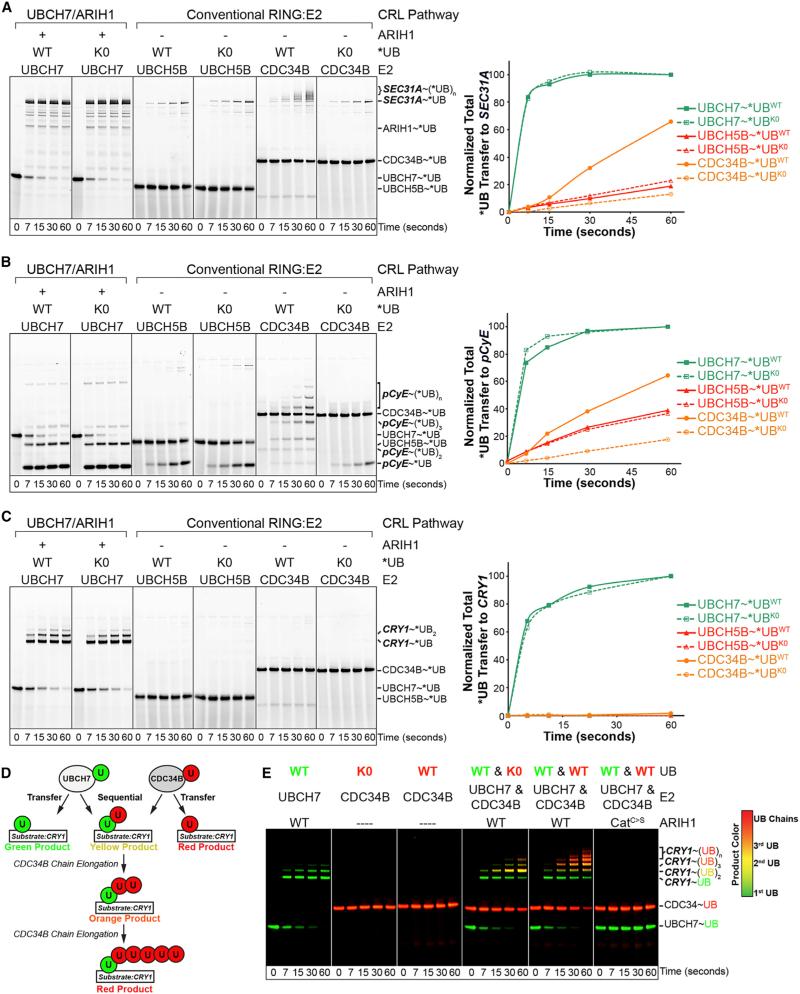
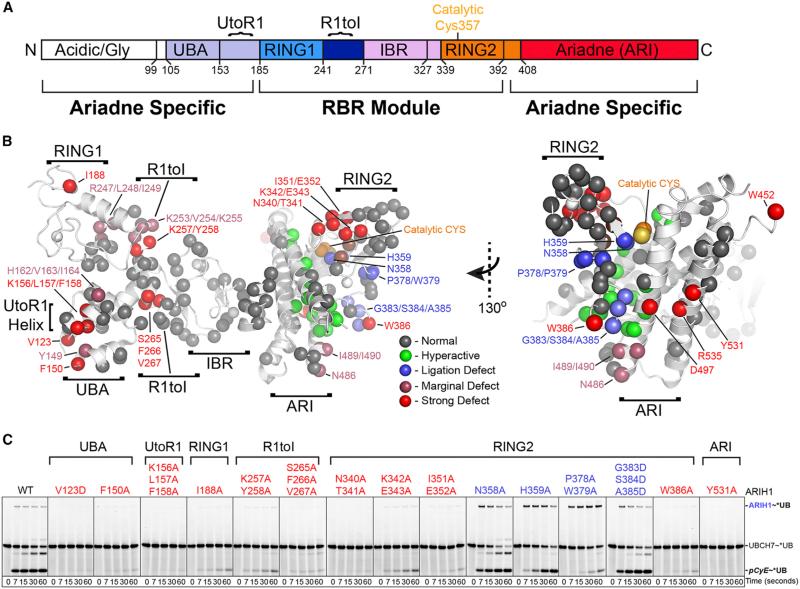
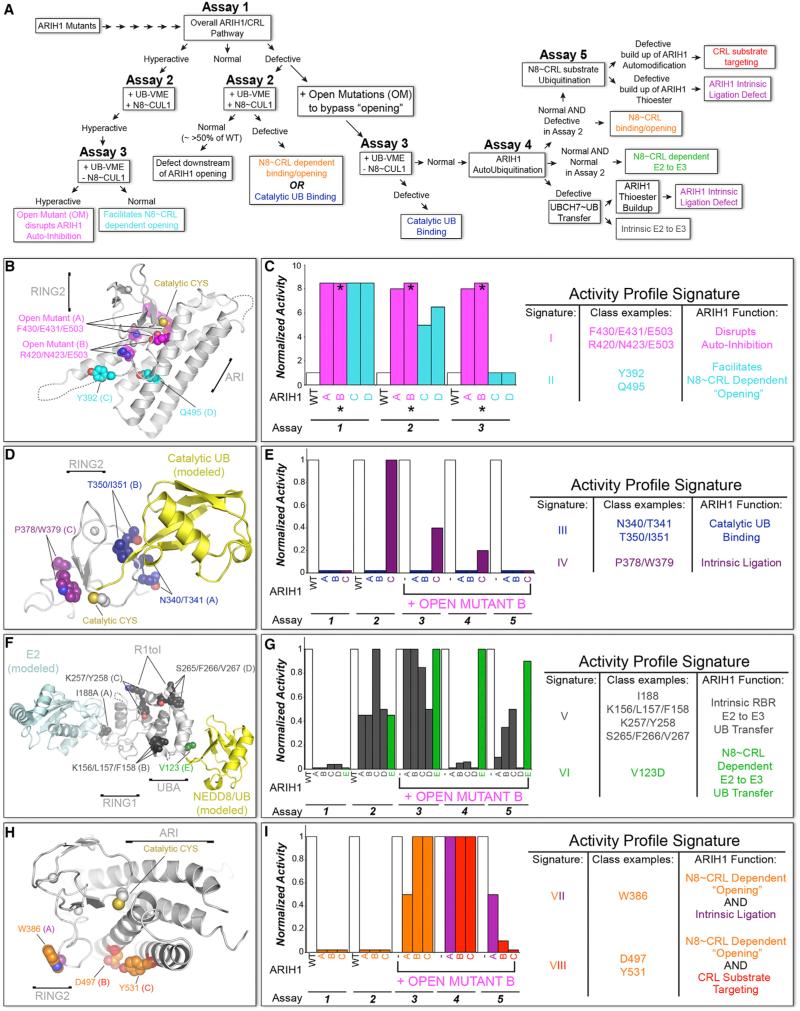
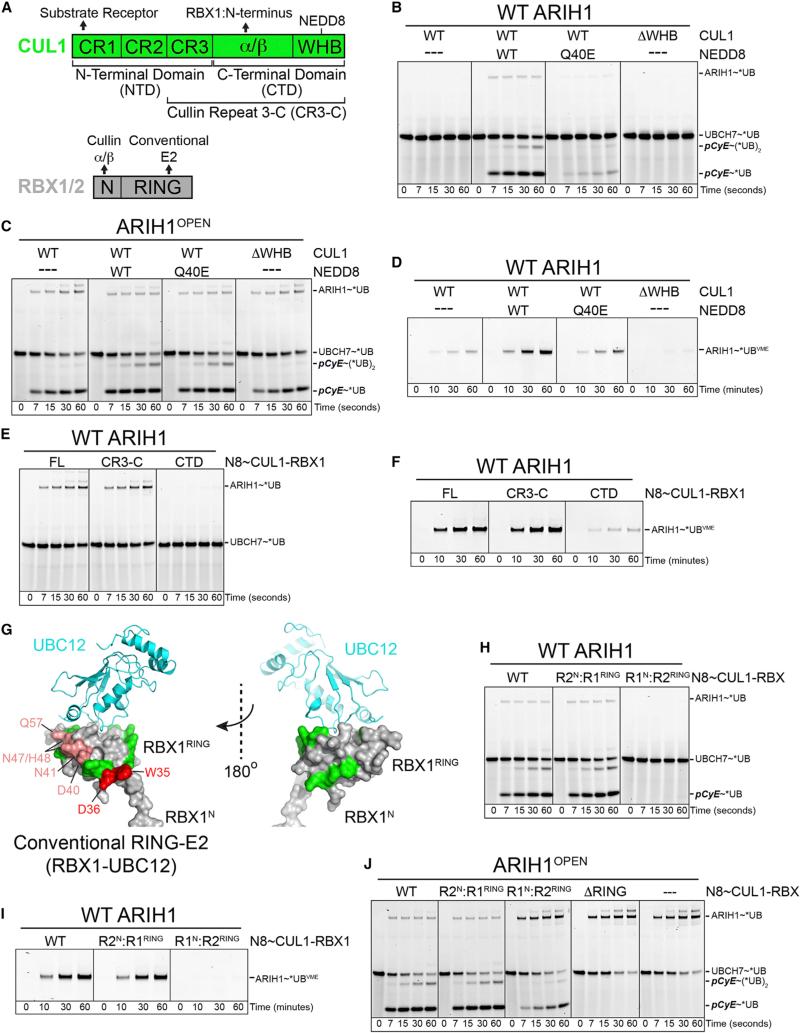
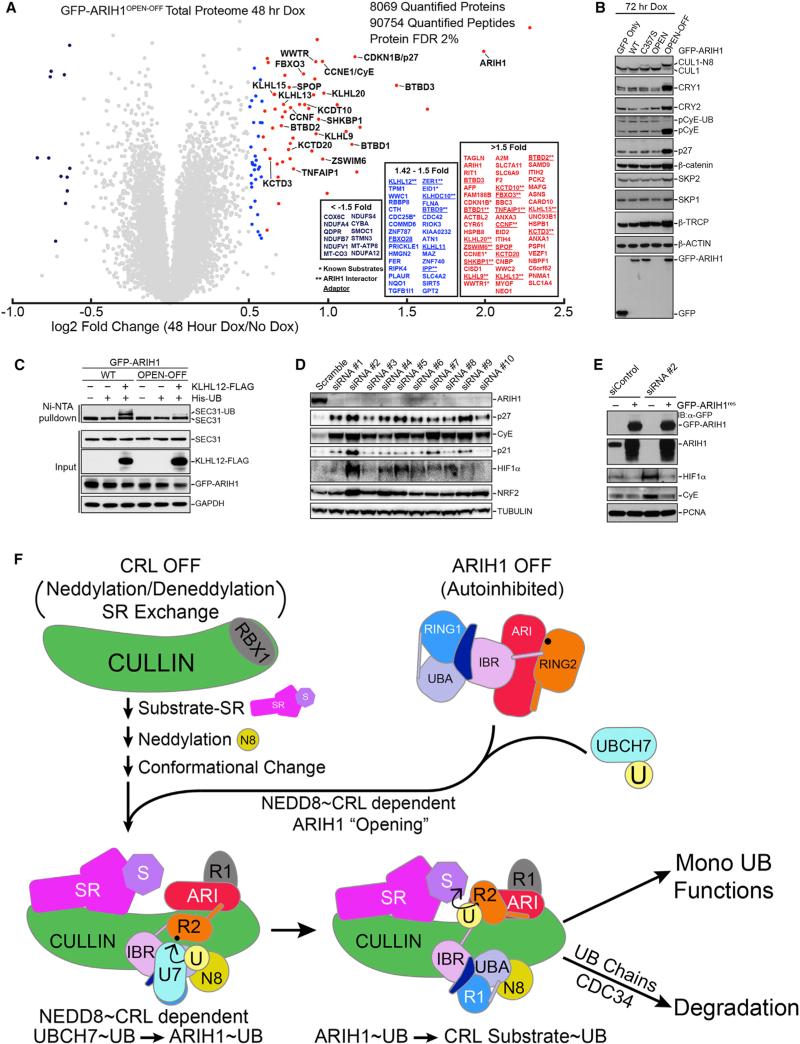
Comment in
-
Tag Team Ubiquitin Ligases.Cell. 2016 Aug 25;166(5):1080-1081. doi: 10.1016/j.cell.2016.08.014. Cell. 2016. PMID: 27565338
References
-
- Berndsen CE, Wolberger C. New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 2014;21:301–307. - PubMed
-
- Busino L, Bassermann F, Maiolica A, Lee C, Nolan PM, Godinho SI, Draetta GF, Pagano M. SCFFbxl3 controls the oscillation of the circadian clock by directing the degradation of cryptochrome proteins. Science. 2007;316:900–904. - PubMed
-
- Ceccarelli DF, Tang X, Pelletier B, Orlicky S, Xie W, Plantevin V, Neculai D, Chou YC, Ogunjimi A, Al-Hakim A, et al. An allosteric inhibitor of the human Cdc34 ubiquitin-conjugating enzyme. Cell. 2011;145:1075–1087. - PubMed
-
- Deshaies RJ, Joazeiro CA. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009;78:399–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
