

Chlorobaculum tepidum Modulates Amino Acid Composition in Response to Energy Availability, as Revealed by a Systematic Exploration of the Energy Landscape of Phototrophic Sulfur Oxidation
- PMID: 27565613
- PMCID: PMC5066360
- DOI: 10.1128/AEM.02111-16
Chlorobaculum tepidum Modulates Amino Acid Composition in Response to Energy Availability, as Revealed by a Systematic Exploration of the Energy Landscape of Phototrophic Sulfur Oxidation
Abstract
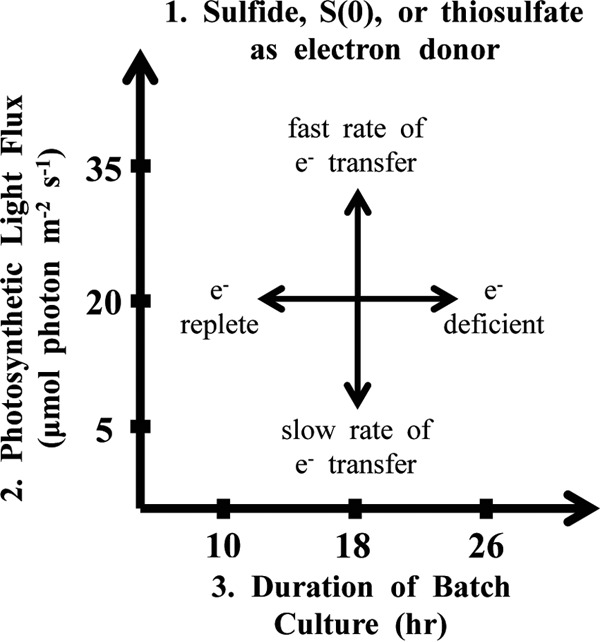
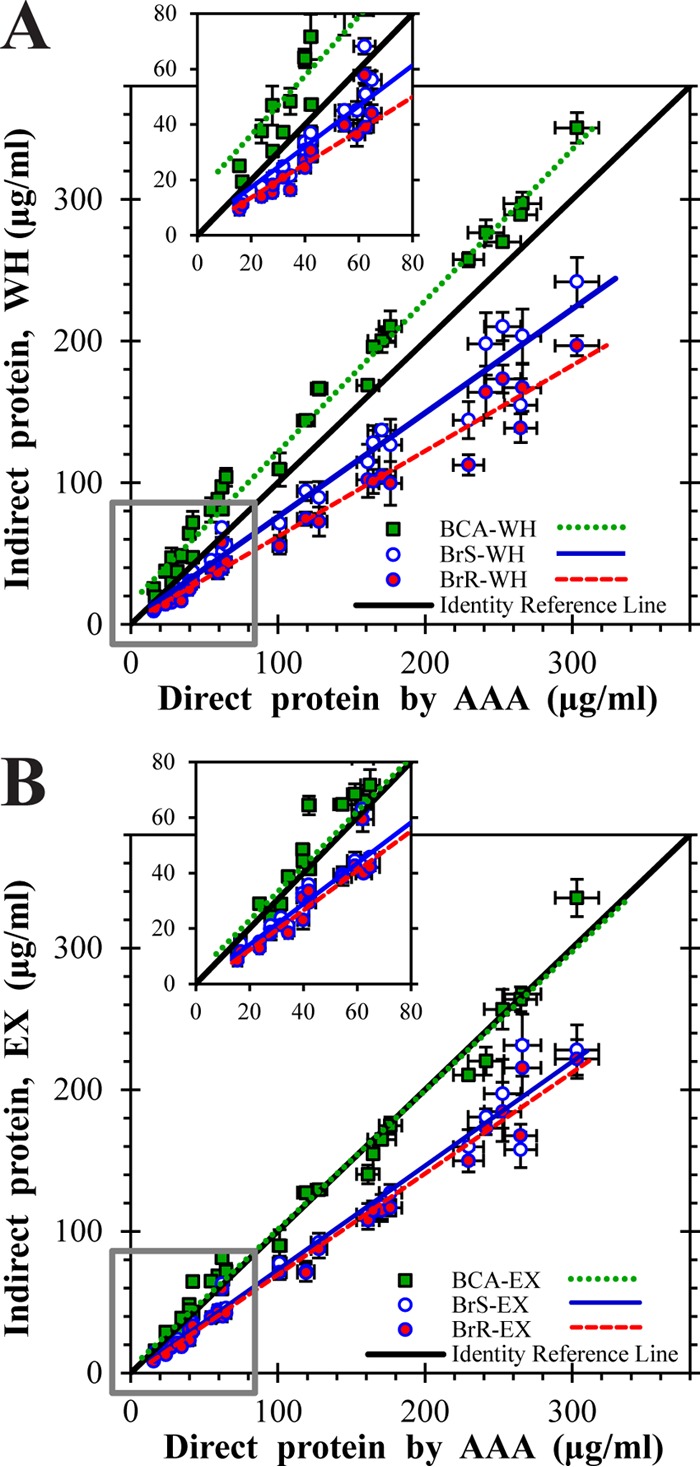
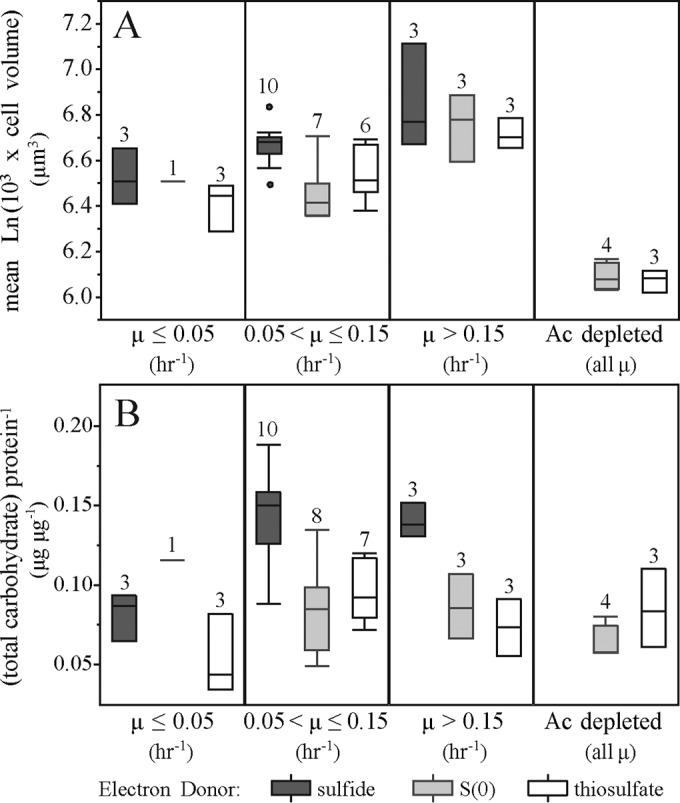
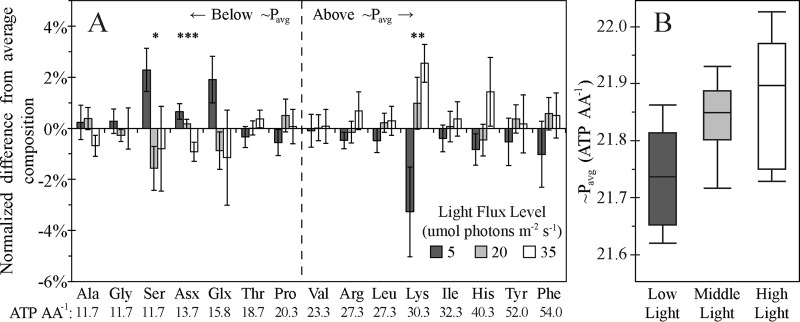
Microbial sulfur metabolism, particularly the formation and consumption of insoluble elemental sulfur (S0), is an important biogeochemical engine that has been harnessed for applications ranging from bioleaching and biomining to remediation of waste streams. Chlorobaculum tepidum, a low-light-adapted photoautolithotrophic sulfur-oxidizing bacterium, oxidizes multiple sulfur species and displays a preference for more reduced electron donors: sulfide > S0 > thiosulfate. To understand this preference in the context of light energy availability, an "energy landscape" of phototrophic sulfur oxidation was constructed by varying electron donor identity, light flux, and culture duration. Biomass and cellular parameters of C. tepidum cultures grown across this landscape were analyzed. From these data, a correction factor for colorimetric protein assays was developed, enabling more accurate biomass measurements for C. tepidum, as well as other organisms. C. tepidum's bulk amino acid composition correlated with energy landscape parameters, including a tendency toward less energetically expensive amino acids under reduced light flux. This correlation, paired with an observation of increased cell size and storage carbon production under electron-rich growth conditions, suggests that C. tepidum has evolved to cope with changing energy availability by tuning its proteome for energetic efficiency and storing compounds for leaner times.
Importance: How microbes cope with and adapt to varying energy availability is an important factor in understanding microbial ecology and in designing efficient biotechnological processes. We explored the response of a model phototrophic organism, Chlorobaculum tepidum, across a factorial experimental design that enabled simultaneous variation and analysis of multiple growth conditions, what we term the "energy landscape." C. tepidum biomass composition shifted toward less energetically expensive amino acids at low light levels. This observation provides experimental evidence for evolved efficiencies in microbial proteomes and emphasizes the role that energy flux may play in the adaptive responses of organisms. From a practical standpoint, our data suggest that bulk biomass amino acid composition could provide a simple proxy to monitor and identify energy stress in microbial systems.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Differential RNA Sequencing Implicates Sulfide as the Master Regulator of S0 Metabolism in Chlorobaculum tepidum and Other Green Sulfur Bacteria.Appl Environ Microbiol. 2018 Jan 17;84(3):e01966-17. doi: 10.1128/AEM.01966-17. Print 2018 Feb 1. Appl Environ Microbiol. 2018. PMID: 29150516 Free PMC article.
-
Quantitative proteomics of Chlorobaculum tepidum: insights into the sulfur metabolism of a phototrophic green sulfur bacterium.FEMS Microbiol Lett. 2011 Oct;323(2):142-50. doi: 10.1111/j.1574-6968.2011.02370.x. Epub 2011 Aug 25. FEMS Microbiol Lett. 2011. PMID: 22092713
-
Chlorobaculum tepidum growth on biogenic S(0) as the sole photosynthetic electron donor.Environ Microbiol. 2016 Sep;18(9):2856-67. doi: 10.1111/1462-2920.12995. Epub 2015 Aug 28. Environ Microbiol. 2016. PMID: 26234460
-
Inorganic sulfur oxidizing system in green sulfur bacteria.Photosynth Res. 2010 Jun;104(2-3):163-76. doi: 10.1007/s11120-010-9531-2. Epub 2010 Feb 9. Photosynth Res. 2010. PMID: 20143161 Review.
-
Sulfur metabolism in phototrophic sulfur bacteria.Adv Microb Physiol. 2009;54:103-200. doi: 10.1016/S0065-2911(08)00002-7. Adv Microb Physiol. 2009. PMID: 18929068 Review.
Cited by
-
Differential RNA Sequencing Implicates Sulfide as the Master Regulator of S0 Metabolism in Chlorobaculum tepidum and Other Green Sulfur Bacteria.Appl Environ Microbiol. 2018 Jan 17;84(3):e01966-17. doi: 10.1128/AEM.01966-17. Print 2018 Feb 1. Appl Environ Microbiol. 2018. PMID: 29150516 Free PMC article.
References
-
- Fortuny M, Guisasola A, Casas C, Gamisans X, Lafuente J, Gabriel D. 2010. Oxidation of biologically produced elemental sulfur under neutrophilic conditions. J Chem Technol Biotechnol 85:378–386. doi:10.1002/jctb.2333. - DOI
-
- Janssen AJH, Ruitenberg R, Buisman CJN. 2001. Industrial applications of new sulphur biotechnology. Water Sci Technol 44:85–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
