

mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach
- PMID: 27565953
- PMCID: PMC5316926
- DOI: 10.1111/pbi.12631
mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach
Abstract
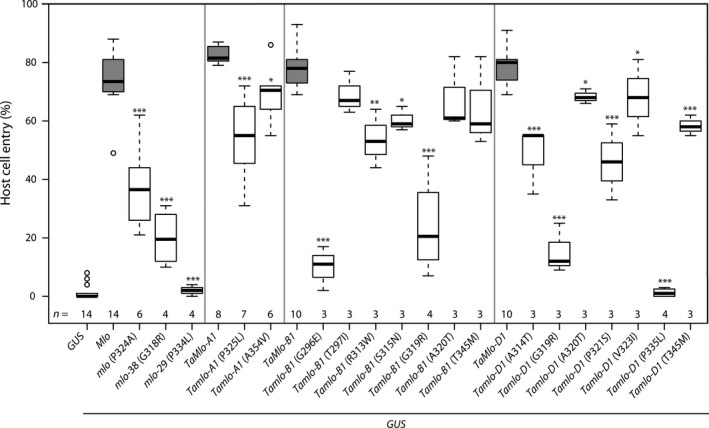
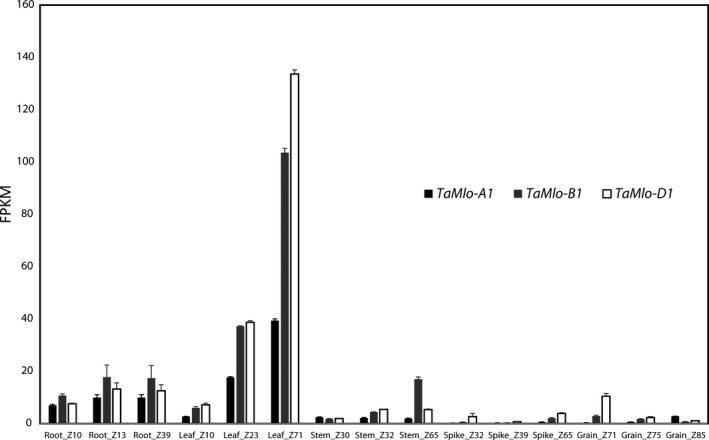
Wheat is one of the most widely grown cereal crops in the world and is an important food grain source for humans. However, wheat yields can be reduced by many abiotic and biotic stress factors, including powdery mildew disease caused by Blumeria graminis f.sp. tritici (Bgt). Generating resistant varieties is thus a major effort in plant breeding. Here, we took advantage of the non-transgenic Targeting Induced Lesions IN Genomes (TILLING) technology to select partial loss-of-function alleles of TaMlo, the orthologue of the barley Mlo (Mildew resistance locus o) gene. Natural and induced loss-of-function alleles (mlo) of barley Mlo are known to confer durable broad-spectrum powdery mildew resistance, typically at the expense of pleiotropic phenotypes such as premature leaf senescence. We identified 16 missense mutations in the three wheat TaMlo homoeologues, TaMlo-A1, TaMlo-B1 and TaMlo-D1 that each lead to single amino acid exchanges. Using transient gene expression assays in barley single cells, we functionally analysed the different missense mutants and identified the most promising candidates affecting powdery mildew susceptibility. By stacking of selected mutant alleles we generated four independent lines with non-conservative mutations in each of the three TaMlo homoeologues. Homozygous triple mutant lines and surprisingly also some of the homozygous double mutant lines showed enhanced, yet incomplete, Bgt resistance without the occurrence of discernible pleiotropic phenotypes. These lines thus represent an important step towards the production of commercial non-transgenic, powdery mildew-resistant bread wheat varieties.
Keywords: Blumeria graminis; Mlo; Targeting Induced Local Lesions in Genomes; hexaploid bread wheat; plant disease resistance; powdery mildew.
© 2016 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures



References
-
- Acevedo‐Garcia, J. , Kusch, S. and Panstruga, R. (2014) Magical mystery tour: MLO proteins in plant immunity and beyond. New Phytol. 204, 273–281. - PubMed
-
- Borrill, P. , Adamski, N. and Uauy, C. (2015) Genomics as the key to unlocking the polyploid potential of wheat. New Phytol. 208, 1008–1022. - PubMed
-
- Brown, J.K.M. and Rant, J.C. (2013) Fitness costs and trade‐offs of disease resistance and their consequences for breeding arable crops. Plant. Pathol. 62, 83–95.
-
- Büschges, R. , Hollricher, K. , Panstruga, R. , Simons, G. , Wolter, M. , Frijters, A. , van Daelen, R. et al. (1997) The barley Mlo gene: a novel control element of plant pathogen resistance. Cell, 88, 695–705. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
