

Hypocretin Neurotransmission Within the Central Amygdala Mediates Escalated Cocaine Self-administration and Stress-Induced Reinstatement in Rats
- PMID: 27567312
- PMCID: PMC5161745
- DOI: 10.1016/j.biopsych.2016.06.010
Hypocretin Neurotransmission Within the Central Amygdala Mediates Escalated Cocaine Self-administration and Stress-Induced Reinstatement in Rats
Abstract
Background: Cocaine addiction is characterized by patterns of compulsive drug-taking, including preoccupation with obtaining cocaine and loss of control over drug intake. The lateral hypothalamic hypocretin/orexin (HCRT) system has been implicated in drug-taking and the reinstatement of drug-seeking. Evidence suggests that HCRT may drive drug-seeking through activation of specific brain regions implicated in stress system dysfunction, including the central amygdala (CeA). The role of HCRT in the persistence of compulsive-like cocaine-taking has yet to be fully elucidated.
Methods: Systemic and intra-CeA microinfusions of the HCRT-receptor 1 antagonist, SB-334867, were administered to rats allowed either short (1 hour; ShA) or long (6 hours; LgA) access to cocaine self-administration. Animals were tested for fixed and progressive ratio responding for cocaine and stress-induced reinstatement of drug-seeking. In addition, using electrophysiological techniques on in vitro slices, we investigated gamma-aminobutyric acidergic (GABAergic) neurotransmission in the medial CeA and the sensitivity of GABAergic synapses to modulation of the HCRT system in ShA or LgA rats.
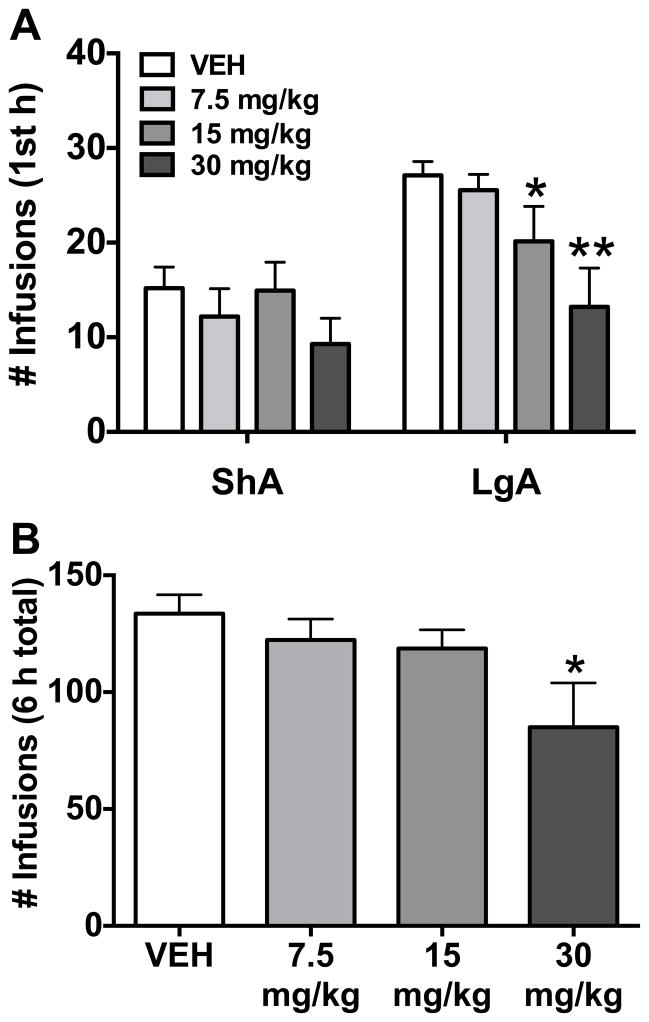
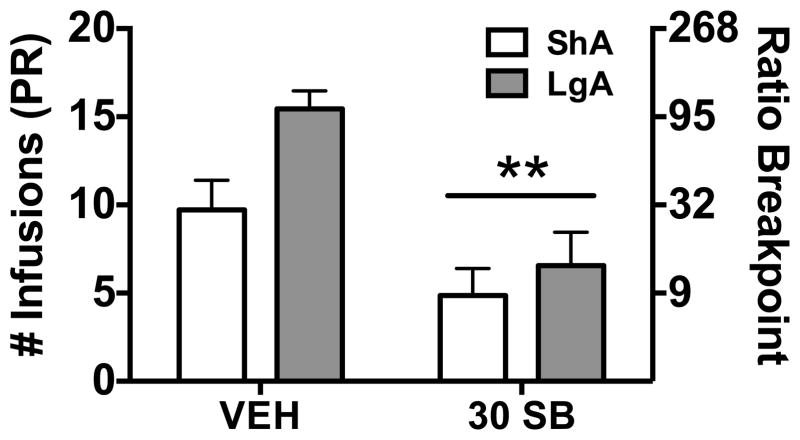
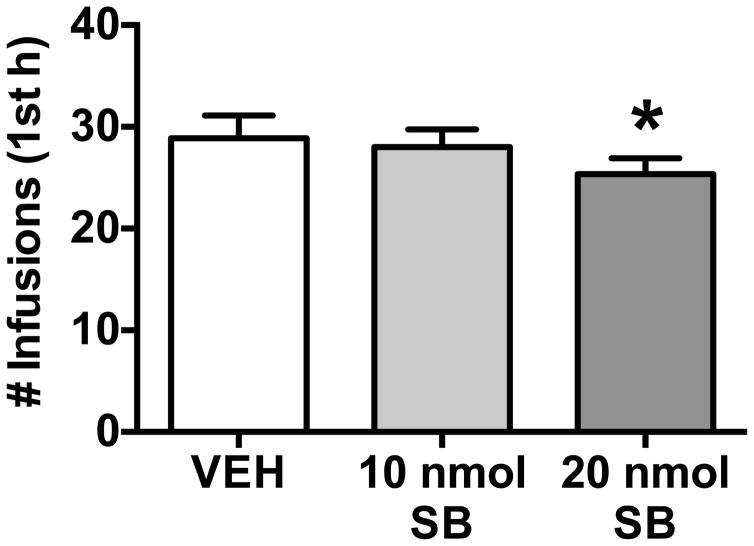
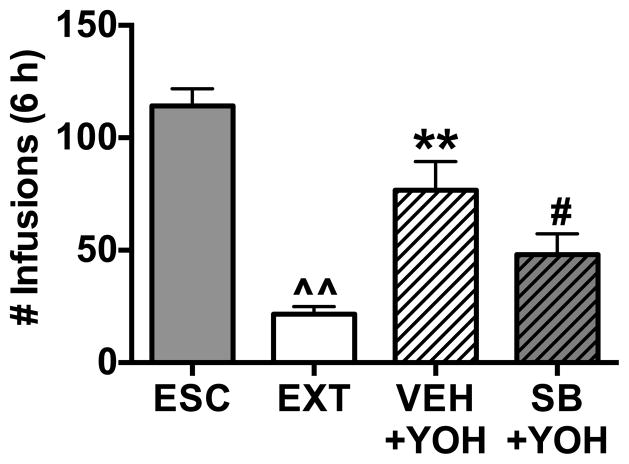
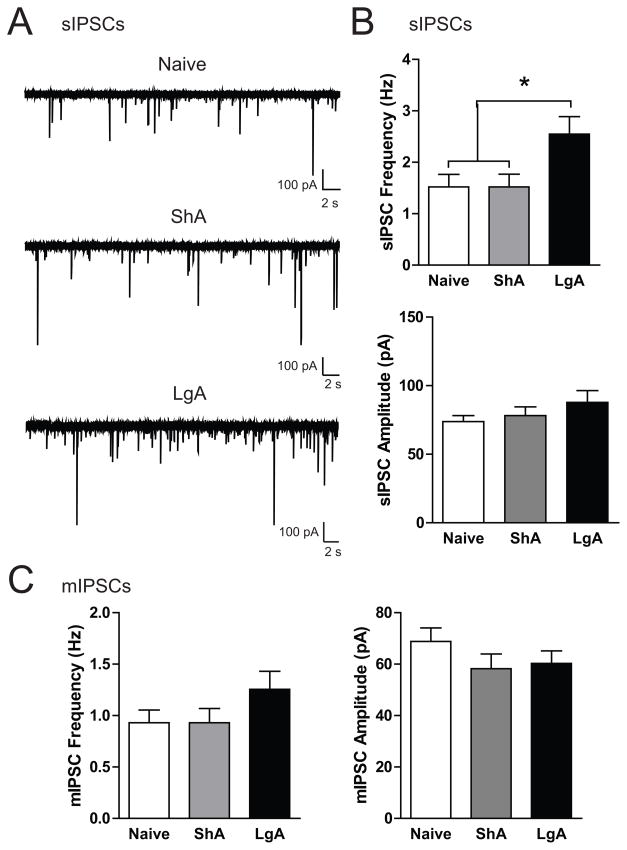
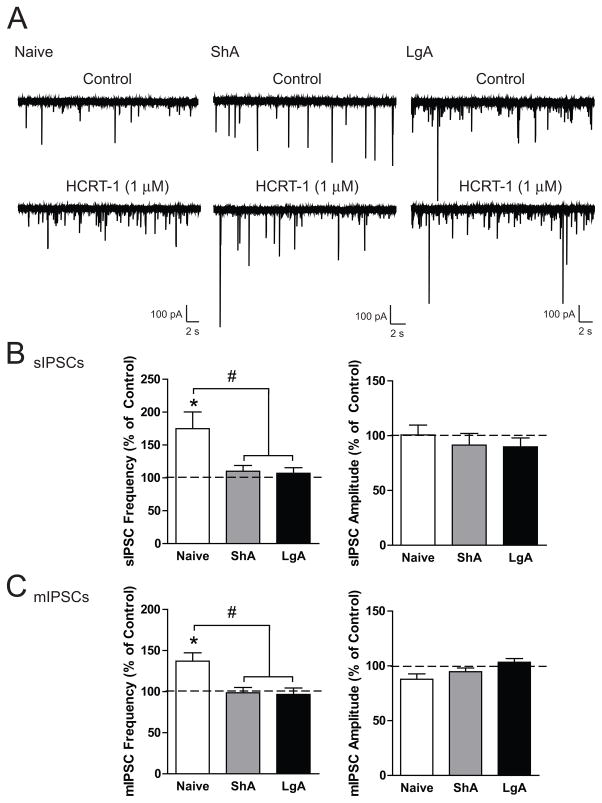
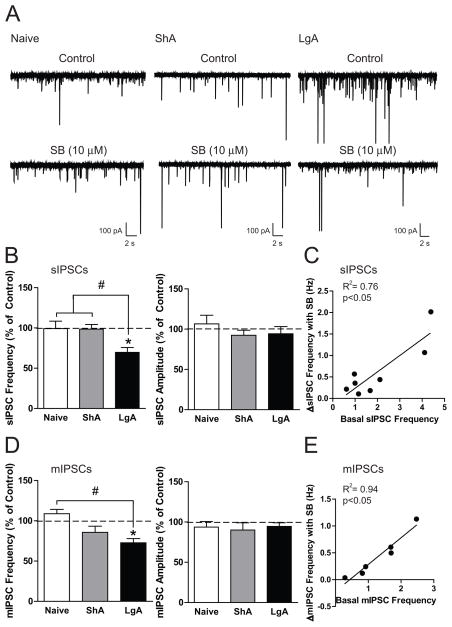
Results: We found systemic administration of SB-334867 (0, 7.5, 15, 30 mg/kg) dose dependently decreased cocaine intake specifically in LgA rats but not in ShA rats. Microinjections of SB-334867 (20 nmol) bilaterally into the CeA significantly reduced cocaine intake in LgA rats. We also observed a significant attenuation of yohimbine-induced reinstatement of cocaine-seeking after intra-CeA SB-334867 (10 nmol) administration. Finally, electrophysiological data indicated enhanced GABAergic neurotransmission within the medial CeA in LgA rats, which was blocked with SB-334867 (10 μmol/L).
Conclusions: These findings suggest that HCRT neurotransmission within the CeA is implicated in compulsive-like cocaine-seeking.
Keywords: Central amygdala; Cocaine; Drug dependence; Hypocretin/orexin; Intravenous self-administration; Reinstatement.
Copyright © 2016 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
Comment in
-
Orexin/Hypocretin, Central Amygdala, and Escalation of Cocaine Intake.Biol Psychiatry. 2017 Apr 1;81(7):552-553. doi: 10.1016/j.biopsych.2017.01.010. Biol Psychiatry. 2017. PMID: 28283054 Free PMC article. No abstract available.
Similar articles
-
Hypocretin receptor 2 antagonism dose-dependently reduces escalated heroin self-administration in rats.Neuropsychopharmacology. 2015 Mar 13;40(5):1123-9. doi: 10.1038/npp.2014.293. Neuropsychopharmacology. 2015. PMID: 25367502 Free PMC article.
-
Kappa opioid receptor-mediated dysregulation of gamma-aminobutyric acidergic transmission in the central amygdala in cocaine addiction.Biol Psychiatry. 2013 Oct 1;74(7):520-8. doi: 10.1016/j.biopsych.2013.04.028. Epub 2013 Jun 14. Biol Psychiatry. 2013. PMID: 23751206 Free PMC article.
-
Orexin-A/Hypocretin-1 Mediates Cocaine-Seeking Behavior in the Posterior Paraventricular Nucleus of the Thalamus via Orexin/Hypocretin Receptor-2.J Pharmacol Exp Ther. 2016 Nov;359(2):273-279. doi: 10.1124/jpet.116.235945. Epub 2016 Aug 18. J Pharmacol Exp Ther. 2016. PMID: 27540003 Free PMC article.
-
Cocaine abuse and midbrain circuits: Functional anatomy of hypocretin/orexin transmission and therapeutic prospect.Brain Res. 2020 Mar 15;1731:146164. doi: 10.1016/j.brainres.2019.02.026. Epub 2019 Feb 20. Brain Res. 2020. PMID: 30796894 Free PMC article. Review.
-
Possible Role of CRF-Hcrt Interaction in the Infralimbic Cortex in the Emergence and Maintenance of Compulsive Alcohol-Seeking Behavior.Alcohol Clin Exp Res. 2020 Feb;44(2):354-367. doi: 10.1111/acer.14264. Epub 2020 Jan 2. Alcohol Clin Exp Res. 2020. PMID: 31840823 Free PMC article. Review.
Cited by
-
Self-Administration of Entactogen Psychostimulants Dysregulates Gamma-Aminobutyric Acid (GABA) and Kappa Opioid Receptor Signaling in the Central Nucleus of the Amygdala of Female Wistar Rats.Front Behav Neurosci. 2021 Dec 16;15:780500. doi: 10.3389/fnbeh.2021.780500. eCollection 2021. Front Behav Neurosci. 2021. PMID: 34975428 Free PMC article.
-
Neurochemical mechanisms and neurocircuitry underlying the contribution of stress to cocaine seeking.J Neurochem. 2021 Jun;157(5):1697-1713. doi: 10.1111/jnc.15340. Epub 2021 Mar 22. J Neurochem. 2021. PMID: 33660857 Free PMC article. Review.
-
The orexin (hypocretin) neuropeptide system is a target for novel therapeutics to treat cocaine use disorder with alcohol coabuse.Neuropharmacology. 2021 Feb 1;183:108359. doi: 10.1016/j.neuropharm.2020.108359. Epub 2020 Oct 19. Neuropharmacology. 2021. PMID: 33091458 Free PMC article. Review.
-
From Molecule to Behavior: Hypocretin/orexin Revisited From a Sex-dependent Perspective.Endocr Rev. 2022 Jul 13;43(4):743-760. doi: 10.1210/endrev/bnab042. Endocr Rev. 2022. PMID: 34792130 Free PMC article. Review.
-
Oxytocin Attenuates Yohimbine-Induced Reinstatement of Alcohol-Seeking in Female Rats via the Central Amygdala.Behav Sci (Basel). 2023 Jul 4;13(7):556. doi: 10.3390/bs13070556. Behav Sci (Basel). 2023. PMID: 37504003 Free PMC article.
References
-
- Berlin GS, Hollander E. Compulsivity, impulsivity, and the DSM-5 process. CNS Spectr. 2014;19:62–68. - PubMed
-
- Pelloux Y, Everitt BJ, Dickinson A. Compulsive drug seeking by rats under punishment: effects of drug taking history. Psychopharmacology. 2007;194:127–137. - PubMed
-
- Ahmed SH, Kenny PJ, Koob GF, Markou A. Neurobiological evidence for hedonic allostasis associated with escalating cocaine use. Nat Neurosci. 2002;5:625–626. - PubMed
-
- Wee S, Specio SE, Koob GF. Effects of dose and session duration on cocaine self-administration in rats. J Pharmacol Exp Ther. 2007;320:1134–1143. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
