

FOXD1-ALDH1A3 Signaling Is a Determinant for the Self-Renewal and Tumorigenicity of Mesenchymal Glioma Stem Cells
- PMID: 27569208
- PMCID: PMC5161538
- DOI: 10.1158/0008-5472.CAN-15-2860
FOXD1-ALDH1A3 Signaling Is a Determinant for the Self-Renewal and Tumorigenicity of Mesenchymal Glioma Stem Cells
Abstract
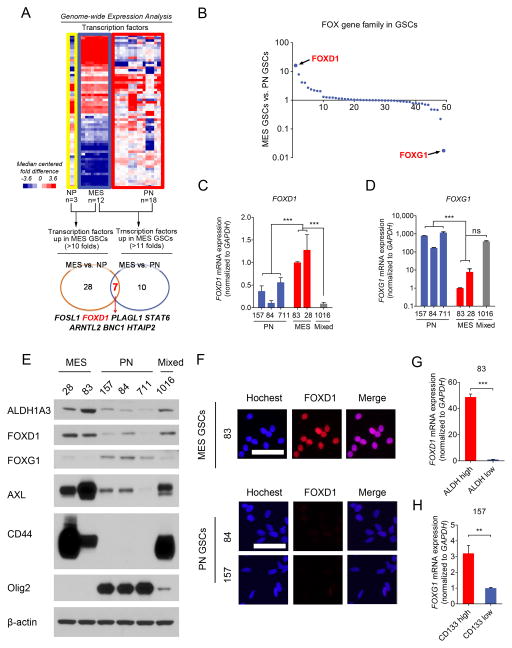
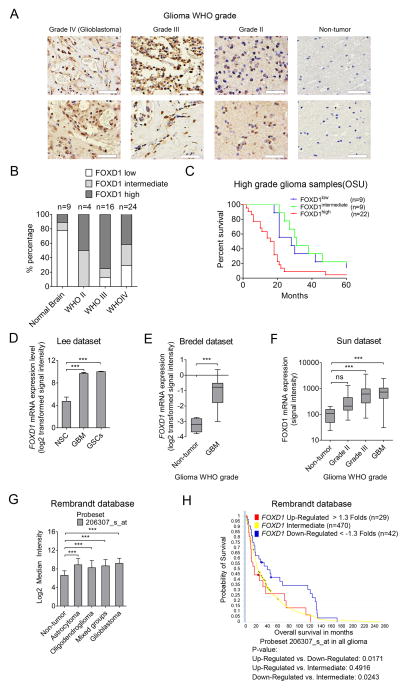
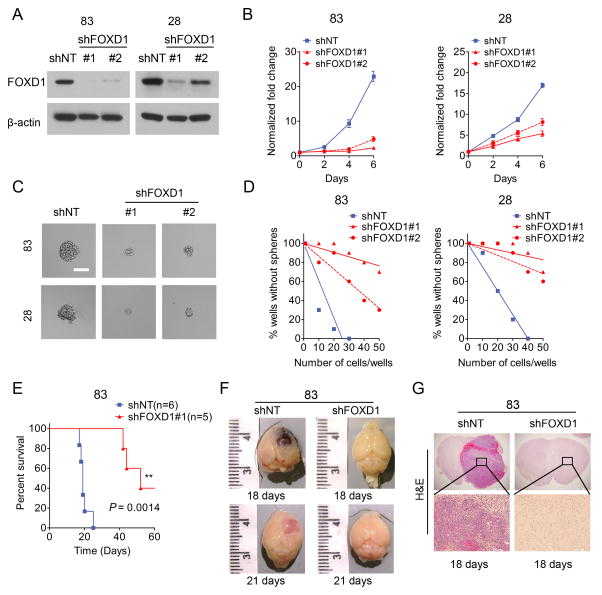
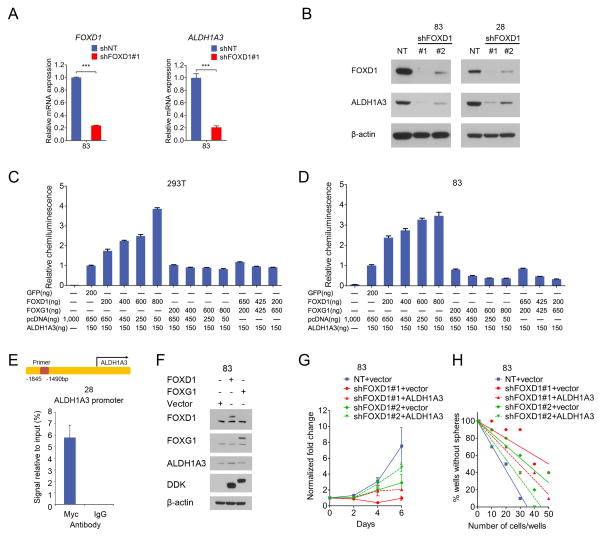
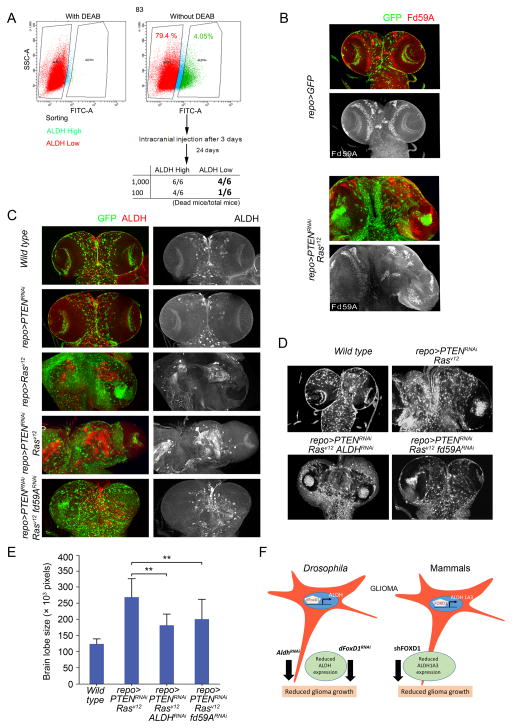
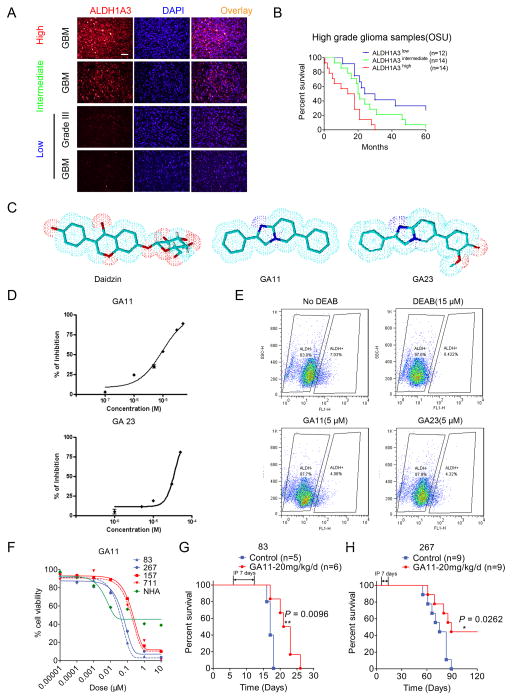
Glioma stem-like cells (GSC) with tumor-initiating activity orchestrate the cellular hierarchy in glioblastoma and engender therapeutic resistance. Recent work has divided GSC into two subtypes with a mesenchymal (MES) GSC population as the more malignant subtype. In this study, we identify the FOXD1-ALDH1A3 signaling axis as a determinant of the MES GSC phenotype. The transcription factor FOXD1 is expressed predominantly in patient-derived cultures enriched with MES, but not with the proneural GSC subtype. shRNA-mediated attenuation of FOXD1 in MES GSC ablates their clonogenicity in vitro and in vivo Mechanistically, FOXD1 regulates the transcriptional activity of ALDH1A3, an established functional marker for MES GSC. Indeed, the functional roles of FOXD1 and ALDH1A3 are likely evolutionally conserved, insofar as RNAi-mediated attenuation of their orthologous genes in Drosophila blocks formation of brain tumors engineered in that species. In clinical specimens of high-grade glioma, the levels of expression of both FOXD1 and ALDH1A3 are inversely correlated with patient prognosis. Finally, a novel small-molecule inhibitor of ALDH we developed, termed GA11, displays potent in vivo efficacy when administered systemically in a murine GSC-derived xenograft model of glioblastoma. Collectively, our findings define a FOXD1-ALDH1A3 pathway in controling the clonogenic and tumorigenic potential of MES GSC in glioblastoma tumors. Cancer Res; 76(24); 7219-30. ©2016 AACR.
©2016 American Association for Cancer Research.
Conflict of interest statement
of Potential Conflicts of Interest No potential conflicts of interest were disclosed by the authors.
Figures
Similar articles
-
The stem cell/cancer stem cell marker ALDH1A3 regulates the expression of the survival factor tissue transglutaminase, in mesenchymal glioma stem cells.Oncotarget. 2017 Apr 4;8(14):22325-22343. doi: 10.18632/oncotarget.16479. Oncotarget. 2017. PMID: 28423611 Free PMC article.
-
USP9X deubiquitinates ALDH1A3 and maintains mesenchymal identity in glioblastoma stem cells.J Clin Invest. 2019 Apr 8;129(5):2043-2055. doi: 10.1172/JCI126414. eCollection 2019 Apr 8. J Clin Invest. 2019. PMID: 30958800 Free PMC article.
-
Mesenchymal glioma stem cells are maintained by activated glycolytic metabolism involving aldehyde dehydrogenase 1A3.Proc Natl Acad Sci U S A. 2013 May 21;110(21):8644-9. doi: 10.1073/pnas.1221478110. Epub 2013 May 6. Proc Natl Acad Sci U S A. 2013. PMID: 23650391 Free PMC article. Clinical Trial.
-
Stem cell signature in glioblastoma: therapeutic development for a moving target.J Neurosurg. 2015 Feb;122(2):324-30. doi: 10.3171/2014.9.JNS132253. Epub 2014 Nov 14. J Neurosurg. 2015. PMID: 25397368 Review.
-
Harnessing OLIG2 function in tumorigenicity and plasticity to target malignant gliomas.Cell Cycle. 2017 Sep 17;16(18):1654-1660. doi: 10.1080/15384101.2017.1361062. Epub 2017 Aug 14. Cell Cycle. 2017. PMID: 28806136 Free PMC article. Review.
Cited by
-
ARL2 overexpression inhibits glioma proliferation and tumorigenicity via down-regulating AXL.BMC Cancer. 2018 May 29;18(1):599. doi: 10.1186/s12885-018-4517-0. BMC Cancer. 2018. PMID: 29843637 Free PMC article.
-
Tumor microenvironment remodeling plus immunotherapy could be used in mesenchymal-like tumor with high tumor residual and drug resistant rate.Commun Biol. 2023 Dec 18;6(1):1281. doi: 10.1038/s42003-023-05667-4. Commun Biol. 2023. PMID: 38110614 Free PMC article.
-
Understanding Glioblastoma Biomarkers: Knocking a Mountain with a Hammer.Cells. 2020 May 16;9(5):1236. doi: 10.3390/cells9051236. Cells. 2020. PMID: 32429463 Free PMC article. Review.
-
FOXD1 is associated with poor outcome and maintains tumor-promoting enhancer-gene programs in basal-like breast cancer.Front Oncol. 2023 May 10;13:1156111. doi: 10.3389/fonc.2023.1156111. eCollection 2023. Front Oncol. 2023. PMID: 37234983 Free PMC article.
-
Mesenchymal Transformation: The Rosetta Stone of Glioblastoma Pathogenesis and Therapy Resistance.Adv Sci (Weinh). 2020 Sep 28;7(22):2002015. doi: 10.1002/advs.202002015. eCollection 2020 Nov. Adv Sci (Weinh). 2020. PMID: 33240762 Free PMC article. Review.
References
-
- Wen PY, Kesari S. Malignant gliomas in adults. The New England journal of medicine. 2008;359:492–507. - PubMed
-
- Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. The New England journal of medicine. 2005;352:987–96. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
