

HMGB1, IL-1α, IL-33 and S100 proteins: dual-function alarmins
- PMID: 27569562
- PMCID: PMC5214941
- DOI: 10.1038/cmi.2016.34
HMGB1, IL-1α, IL-33 and S100 proteins: dual-function alarmins
Abstract
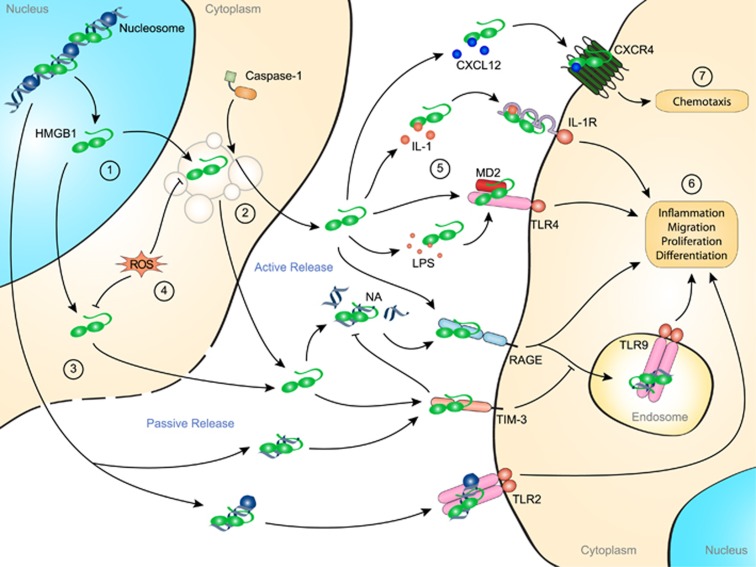
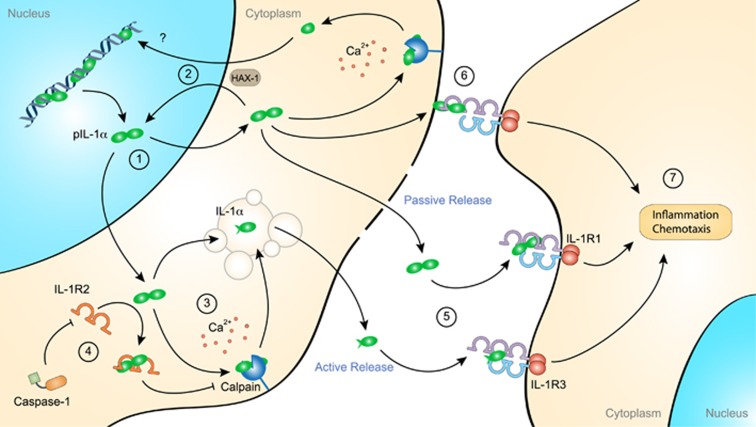
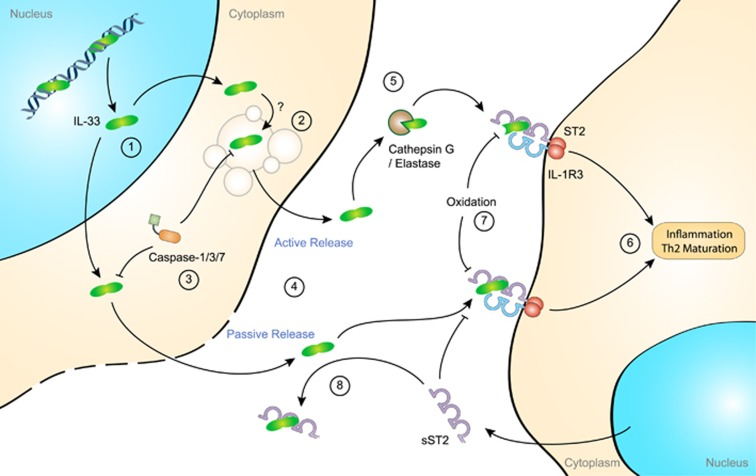
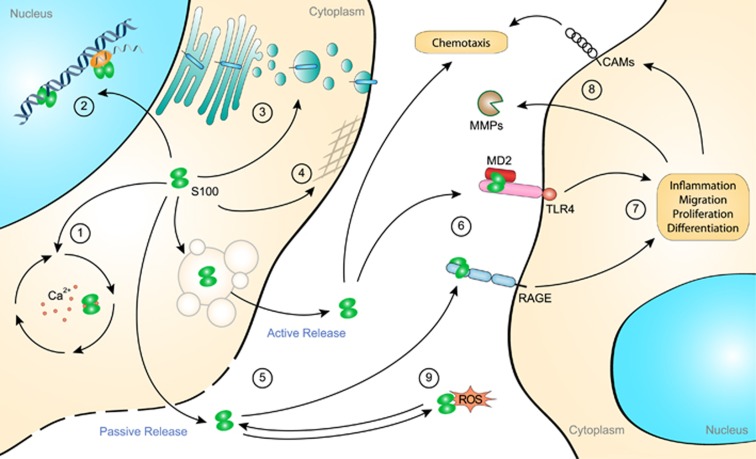
Our immune system is based on the close collaboration of the innate and adaptive immune systems for the rapid detection of any threats to the host. Recognition of pathogen-derived molecules is entrusted to specific germline-encoded signaling receptors. The same receptors have now also emerged as efficient detectors of misplaced or altered self-molecules that signal tissue damage and cell death following, for example, disruption of the blood supply and subsequent hypoxia. Many types of endogenous molecules have been shown to provoke such sterile inflammatory states when released from dying cells. However, a group of proteins referred to as alarmins have both intracellular and extracellular functions which have been the subject of intense research. Indeed, alarmins can either exert beneficial cell housekeeping functions, leading to tissue repair, or provoke deleterious uncontrolled inflammation. This group of proteins includes the high-mobility group box 1 protein (HMGB1), interleukin (IL)-1α, IL-33 and the Ca2+-binding S100 proteins. These dual-function proteins share conserved regulatory mechanisms, such as secretory routes, post-translational modifications and enzymatic processing, that govern their extracellular functions in time and space. Release of alarmins from mesenchymal cells is a highly relevant mechanism by which immune cells can be alerted of tissue damage, and alarmins play a key role in the development of acute or chronic inflammatory diseases and in cancer development.
Figures
References
-
- Janeway CA. Pillars article: approaching the asymptote? Evolution and revolution in immunology. Cold spring harb symp quant boil. 1989. 54: 1–13. J Immunol 2013; 191: 4475–4487. - PubMed
-
- Matzinger P. Tolerance, danger, and the extended family. Annu Rev Immunol 1994; 12: 991–1045. - PubMed
-
- Seong S-Y, Matzinger P. Hydrophobicity: an ancient damage-associated molecular pattern that initiates innate immune responses. Nat Rev Immunol 2004; 4: 469–478. - PubMed
-
- Bessa J, Meyer CA, de Vera Mudry MC, Schlicht S, Smith SH, Iglesias A et al. Altered subcellular localization of IL-33 leads to non-resolving lethal inflammation. J Autoimmun 2014; 55: 33–41. - PubMed
-
- Agresti A, Bianchi ME. HMGB proteins and gene expression. Curr Opin Genet Dev 2003; 13: 170–178. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
