

Role of the host restriction factor APOBEC3 on papillomavirus evolution
- PMID: 27570633
- PMCID: PMC4999249
- DOI: 10.1093/ve/vev015
Role of the host restriction factor APOBEC3 on papillomavirus evolution
Abstract
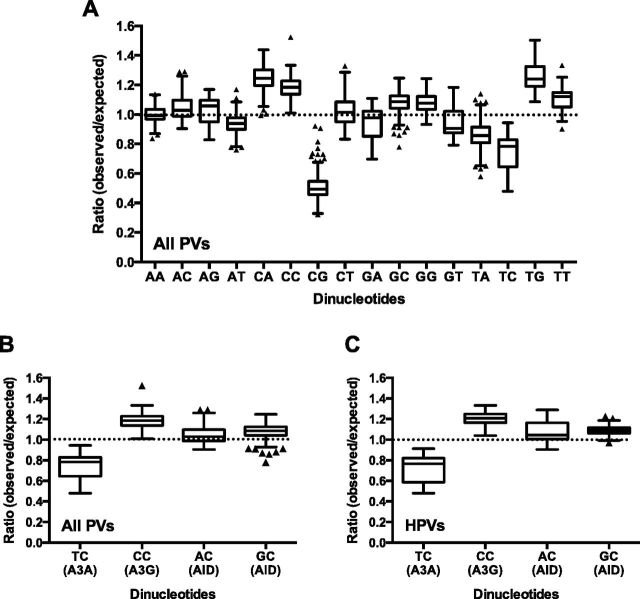
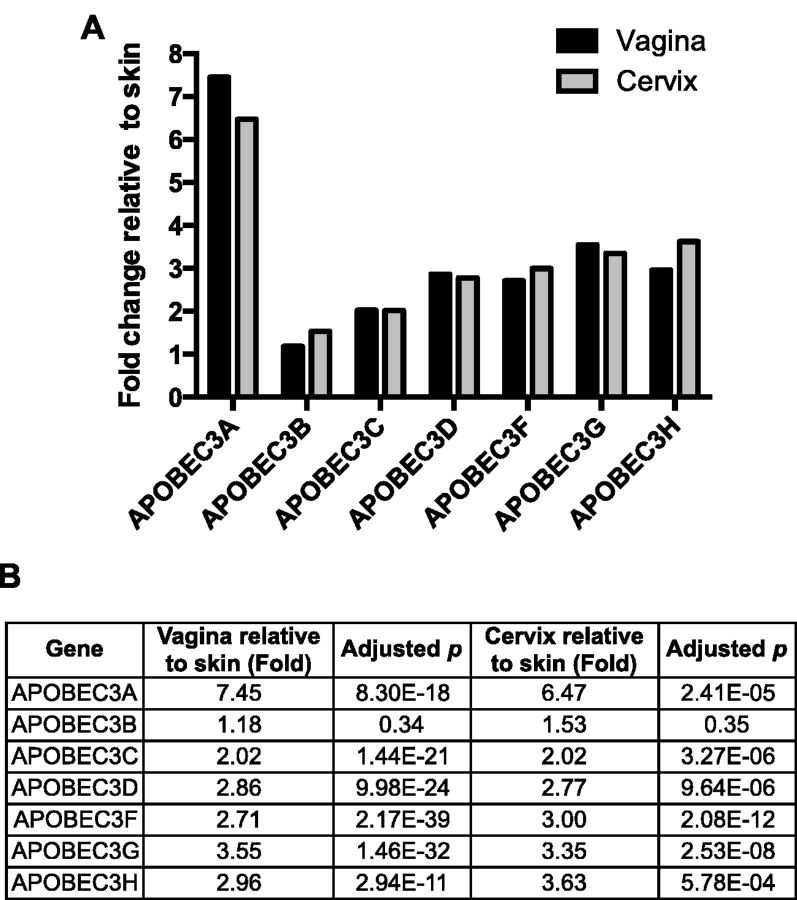
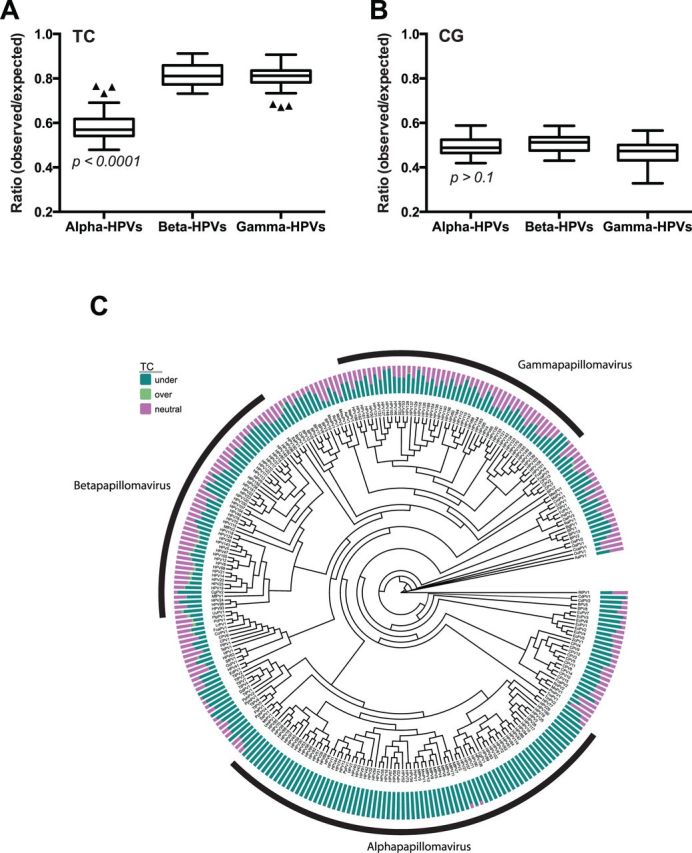
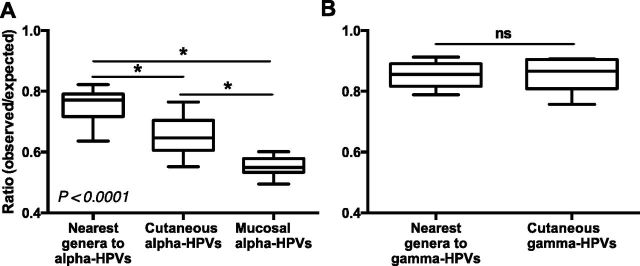
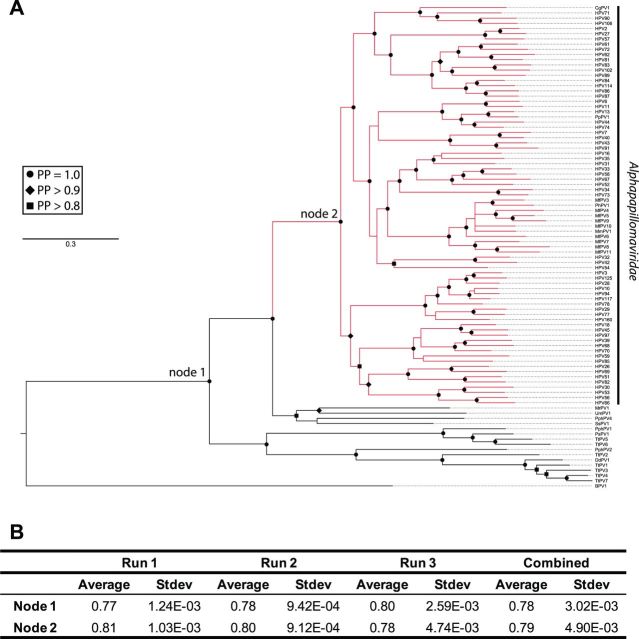
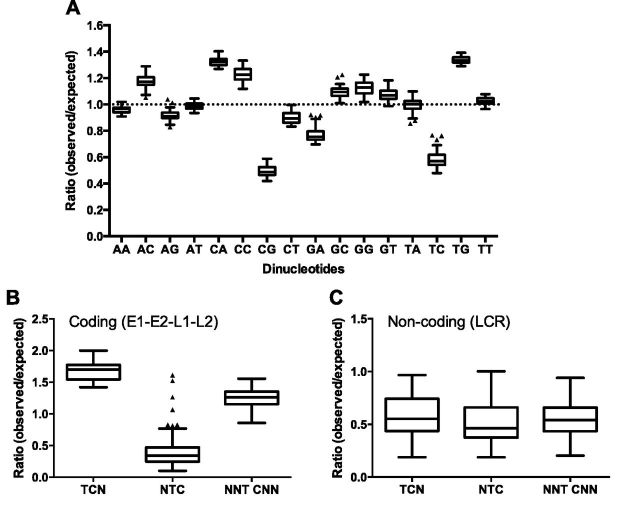
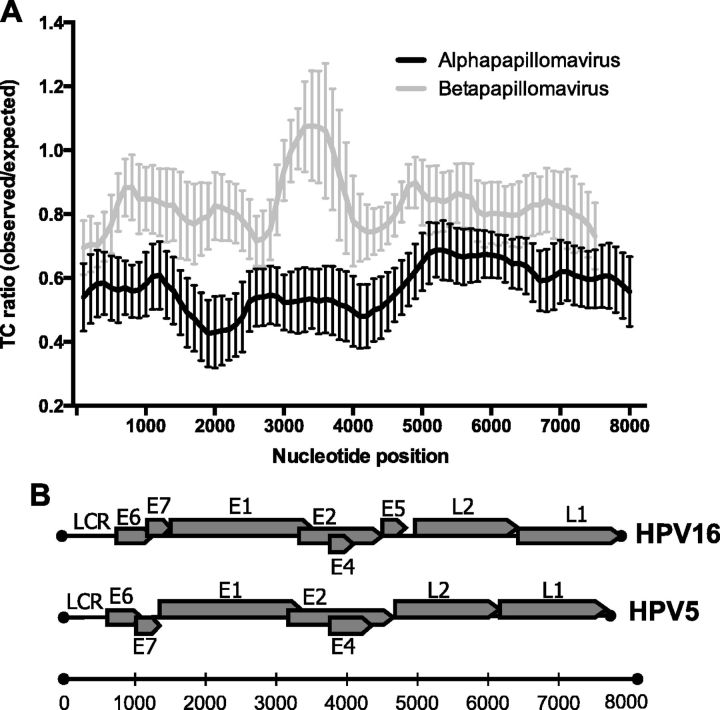
More than 270 different types of papillomaviruses have been discovered in a wide array of animal species. Despite the great diversity of papillomaviruses, little is known about the evolutionary processes that drive host tropism and the emergence of oncogenic genotypes. Although host defense mechanisms have evolved to interfere with various aspects of a virus life cycle, viruses have also coevolved copious strategies to avoid host antiviral restriction. Our and other studies have shown that the cytidine deaminase APOBEC3 family members edit HPV genomes and restrict virus infectivity. Thus, we hypothesized that host restriction by APOBEC3 served as selective pressure during papillomavirus evolution. To test this hypothesis, we analyzed the relative abundance of all dinucleotide sequences in full-length genomes of 274 papillomavirus types documented in the Papillomavirus Episteme database (PaVE). Here, we report that TC dinucleotides, the preferred target sequence of several human APOBEC3 proteins (hA3A, hA3B, hA3F, and hA3H), are highly depleted in papillomavirus genomes. Given that HPV infection is highly tissue-specific, the expression levels of APOBEC3 family members were analyzed. The basal expression levels of all APOBEC3 isoforms, excluding hA3B, are significantly higher in mucosal skin compared with cutaneous skin. Interestingly, we reveal that Alphapapillomaviruses (alpha-PVs), a majority of which infects anogenital mucosa, display the most dramatic reduction in TC dinucleotide content. Computer modeling and reconstruction of ancestral alpha-PV genomes suggest that TC depletion occurred after the alpha-PVs diverged from their most recent common ancestor. In addition, we found that TC depletion in alpha-PVs is greatly affected by protein coding potential. Taken together, our results suggest that PVs replicating in tissues with high APOBEC3 levels may have evolved to evade restriction by selecting for variants that contain reduced APOBEC3 target sites in their genomes.
Keywords: APOBEC3; coevolution; papillomavirus; restriction factor.
Conflict of interest statement
Conflicts of Interest: None declared
Figures
References
-
- Ahasan M. M., et al. (2015). ‘APOBEC3A and C Decrease Human Papillomavirus 16 Pseudovirion Infectivity’, Biochemical and Biophysical Research Communications, 457: 295–9. - PubMed
-
- Barker D., Meade A., Pagel M. (2007). ‘Constrained Models of Evolution Lead to Improved Prediction of Functional Linkage from Correlated Gain and Loss of Genes’, Bioinformatics, 23: 14–20. - PubMed
-
- Betts M. J., Russell R. B. (2003). ‘Amino Acid Properties and Consequences of Substitutions’, Bioinformatics for Geneticists, 317: 289.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
