

Repetition suppression: a means to index neural representations using BOLD?
- PMID: 27574308
- PMCID: PMC5003856
- DOI: 10.1098/rstb.2015.0355
Repetition suppression: a means to index neural representations using BOLD?
Abstract
Understanding how the human brain gives rise to complex cognitive processes remains one of the biggest challenges of contemporary neuroscience. While invasive recording in animal models can provide insight into neural processes that are conserved across species, our understanding of cognition more broadly relies upon investigation of the human brain itself. There is therefore an imperative to establish non-invasive tools that allow human brain activity to be measured at high spatial and temporal resolution. In recent years, various attempts have been made to refine the coarse signal available in functional magnetic resonance imaging (fMRI), providing a means to investigate neural activity at the meso-scale, i.e. at the level of neural populations. The most widely used techniques include repetition suppression and multivariate pattern analysis. Human neuroscience can now use these techniques to investigate how representations are encoded across neural populations and transformed by relevant computations. Here, we review the physiological basis, applications and limitations of fMRI repetition suppression with a brief comparison to multivariate techniques. By doing so, we show how fMRI repetition suppression holds promise as a tool to reveal complex neural mechanisms that underlie human cognitive function.This article is part of the themed issue 'Interpreting BOLD: a dialogue between cognitive and cellular neuroscience'.
Keywords: functional magnetic resonance imaging adaptation; neural computation; neural representation; repetition suppression.
© 2016 The Author(s).
Figures
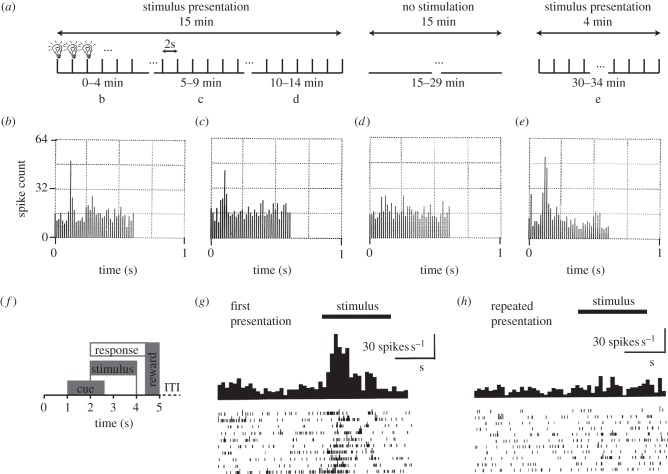
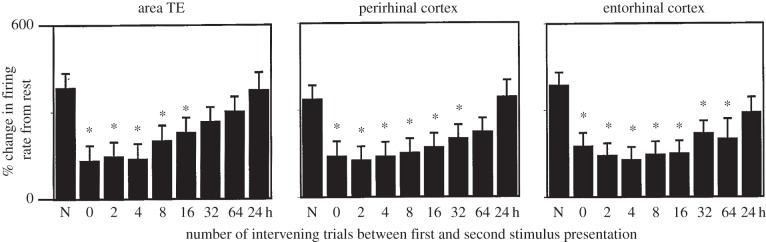
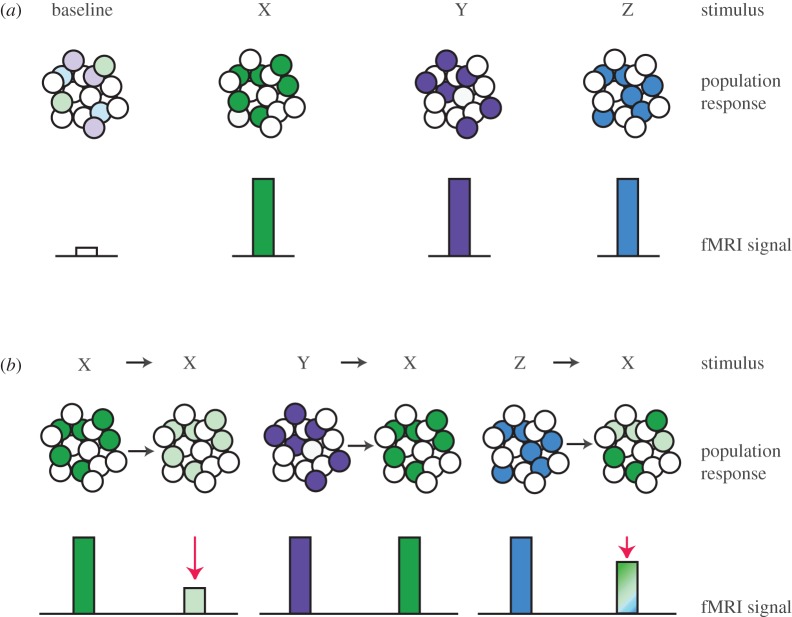
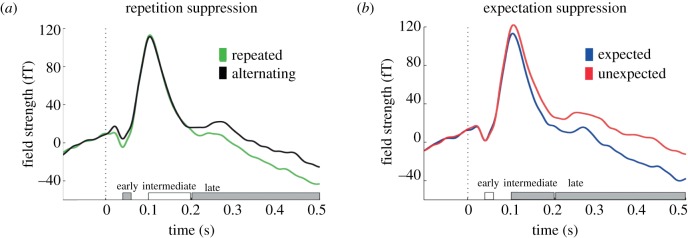
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical