

Conservation and diversity in the ultralong third heavy-chain complementarity-determining region of bovine antibodies
- PMID: 27574710
- PMCID: PMC5000368
- DOI: 10.1126/sciimmunol.aaf7962
Conservation and diversity in the ultralong third heavy-chain complementarity-determining region of bovine antibodies
Abstract
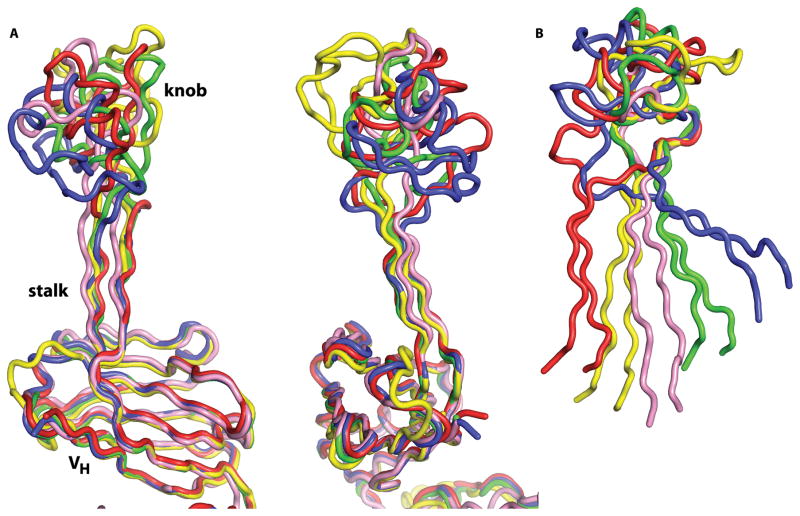
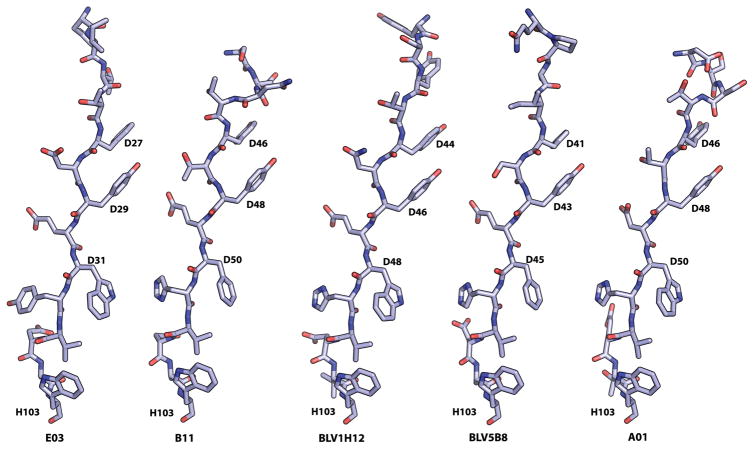
A subset of bovine antibodies have an exceptionally long third heavy-chain complementarity determining region (CDR H3) that is highly variable in sequence and includes multiple cysteines. These long CDR H3s (up to 69 residues) fold into a long stalk atop which sits a knob domain that is located far from the antibody surface. Three new bovine Fab crystal structures have been determined to decipher the conserved and variable features of ultralong CDR H3s that lead to diversity in antigen recognition. Despite high sequence variability, the stalks adopt a conserved β-ribbon structure, while the knob regions share a conserved β-sheet that serves as a scaffold for two connecting loops of variable length and conformation, as well as one conserved disulfide. Variation in patterns and connectivity of the remaining disulfides contribute to the knob structural diversity. The unusual architecture of these ultralong bovine CDR H3s for generating diversity is unique in adaptive immune systems.
Conflict of interest statement
V.S. has equity in Sevion, Inc., which has interest in commercial development of cow antibodies.
Figures





Similar articles
-
Bos taurus ultralong CDR H3 antibodies.Curr Opin Struct Biol. 2016 Jun;38:62-7. doi: 10.1016/j.sbi.2016.05.004. Epub 2016 Jun 10. Curr Opin Struct Biol. 2016. PMID: 27295423 Free PMC article. Review.
-
The Unusual Genetics and Biochemistry of Bovine Immunoglobulins.Adv Immunol. 2018;137:135-164. doi: 10.1016/bs.ai.2017.12.004. Epub 2018 Feb 9. Adv Immunol. 2018. PMID: 29455846 Free PMC article.
-
Germline-Encoded Positional Cysteine Polymorphisms Enhance Diversity in Antibody Ultralong CDR H3 Regions.J Immunol. 2022 Dec 1;209(11):2141-2148. doi: 10.4049/jimmunol.2200455. J Immunol. 2022. PMID: 36426974 Free PMC article.
-
Reshaping antibody diversity.Cell. 2013 Jun 6;153(6):1379-93. doi: 10.1016/j.cell.2013.04.049. Cell. 2013. PMID: 23746848 Free PMC article.
-
Diversity in the Cow Ultralong CDR H3 Antibody Repertoire.Front Immunol. 2018 Jun 4;9:1262. doi: 10.3389/fimmu.2018.01262. eCollection 2018. Front Immunol. 2018. PMID: 29915599 Free PMC article. Review.
Cited by
-
Bos taurus ultralong CDR H3 antibodies.Curr Opin Struct Biol. 2016 Jun;38:62-7. doi: 10.1016/j.sbi.2016.05.004. Epub 2016 Jun 10. Curr Opin Struct Biol. 2016. PMID: 27295423 Free PMC article. Review.
-
Variations in antibody repertoires correlate with vaccine responses.Genome Res. 2022 Apr;32(4):791-804. doi: 10.1101/gr.276027.121. Epub 2022 Mar 31. Genome Res. 2022. PMID: 35361626 Free PMC article.
-
Contributions of Farm Animals to Immunology.Front Vet Sci. 2018 Dec 6;5:307. doi: 10.3389/fvets.2018.00307. eCollection 2018. Front Vet Sci. 2018. PMID: 30574508 Free PMC article. Review.
-
The smallest functional antibody fragment: Ultralong CDR H3 antibody knob regions potently neutralize SARS-CoV-2.Proc Natl Acad Sci U S A. 2023 Sep 26;120(39):e2303455120. doi: 10.1073/pnas.2303455120. Epub 2023 Sep 18. Proc Natl Acad Sci U S A. 2023. PMID: 37722054 Free PMC article.
-
The proximity of the N- and C- termini of bovine knob domains enable engineering of target specificity into polypeptide chains.MAbs. 2022 Jan-Dec;14(1):2076295. doi: 10.1080/19420862.2022.2076295. MAbs. 2022. PMID: 35634719 Free PMC article.
References
-
- Saini SS, Allore B, Jacobs RM, Kaushik A. Exceptionally long CDR3H region with multiple cysteine residues in functional bovine IgM antibodies. Eur J Immunol. 1999;29:2420–2426. - PubMed
-
- Griffin LM, et al. Analysis of heavy and light chain sequences of conventional camelid antibodies from Camelus dromedarius and Camelus bactrianus species. J Immunol Meth. 2014;405:35–46. - PubMed
-
- Diaz M, Stanfield RL, Greenberg AS, Flajnik MF. Structural analysis, selection, and ontogeny of the shark new antigen receptor (IgNAR): identification of a new locus preferentially expressed in early development. Immunogenetics. 2002;54:501–512. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
