

Genetically Encoded Biosensors Reveal PKA Hyperphosphorylation on the Myofilaments in Rabbit Heart Failure
- PMID: 27576469
- PMCID: PMC5307331
- DOI: 10.1161/CIRCRESAHA.116.308964
Genetically Encoded Biosensors Reveal PKA Hyperphosphorylation on the Myofilaments in Rabbit Heart Failure
Abstract
Rationale: In heart failure, myofilament proteins display abnormal phosphorylation, which contributes to contractile dysfunction. The mechanisms underlying the dysregulation of protein phosphorylation on myofilaments is not clear.
Objective: This study aims to understand the mechanisms underlying altered phosphorylation of myofilament proteins in heart failure.
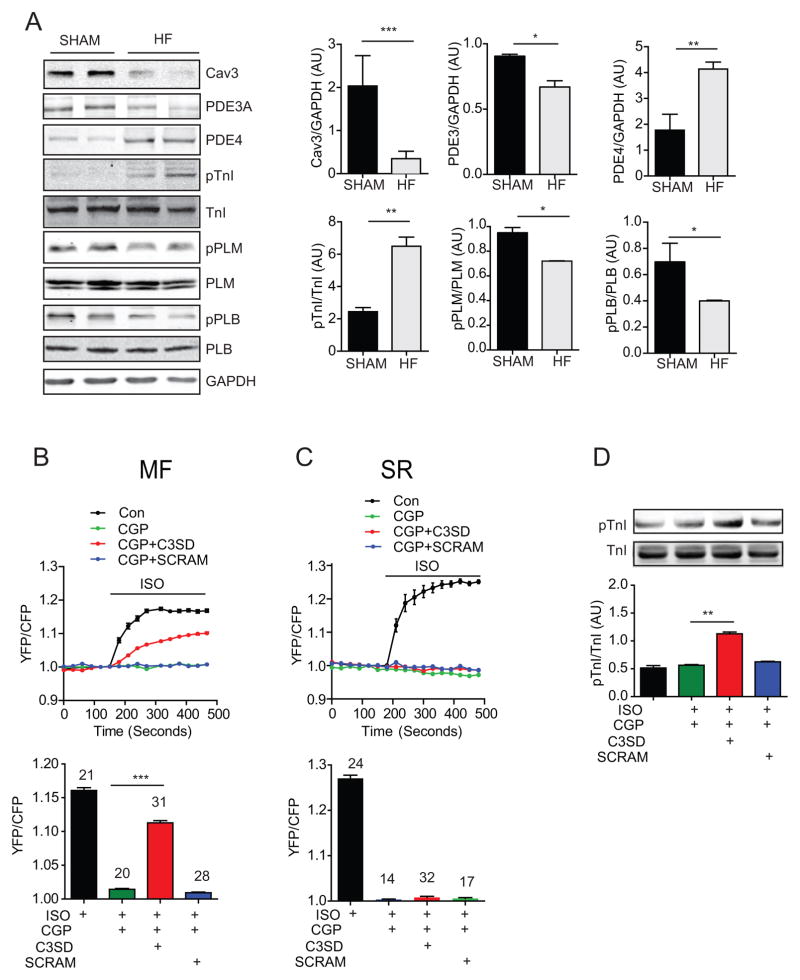
Methods and results: We generate a novel genetically encoded protein kinase A (PKA) biosensor anchored onto the myofilaments in rabbit cardiac myocytes to examine PKA activity at the myofilaments in responses to adrenergic stimulation. We show that PKA activity is shifted from the sarcolemma to the myofilaments in hypertrophic failing rabbit myocytes. In particular, the increased PKA activity on the myofilaments is because of an enhanced β2 adrenergic receptor signal selectively directed to the myofilaments together with a reduced phosphodiesterase activity associated with the myofibrils. Mechanistically, the enhanced PKA activity on the myofilaments is associated with downregulation of caveolin-3 in the hypertrophic failing rabbit myocytes. Reintroduction of caveolin-3 in the failing myocytes is able to normalize the distribution of β2 adrenergic receptor signal by preventing PKA signal access to the myofilaments and to restore contractile response to adrenergic stimulation.
Conclusions: In hypertrophic rabbit myocytes, selectively enhanced β2 adrenergic receptor signaling toward the myofilaments contributes to elevated PKA activity and PKA phosphorylation of myofilament proteins. Reintroduction of caveolin-3 is able to confine β2 adrenergic receptor signaling and restore myocyte contractility in response to β adrenergic stimulation.
Keywords: adrenergic receptor; heart failure; myofibrils; phosphorylation; protein kinase A.
© 2016 American Heart Association, Inc.
Figures







References
-
- Ruehr ML, Russell MA, Bond M. A-kinase anchoring protein targeting of protein kinase A in the heart. J Mol Cell Cardiol. 2004;37:653–65. - PubMed
-
- Koss KL, Kranias EG. Phospholamban: a prominent regulator of myocardial contractility. Circ Res. 1996;79:1059–63. - PubMed
-
- Stelzer JE, Patel JR, Walker JW, Moss RL. Differential roles of cardiac myosin-binding protein C and cardiac troponin I in the myofibrillar force responses to protein kinase A phosphorylation. Circ Res. 2007;101:503–11. - PubMed
-
- Eschenhagen T. Beta-adrenergic signaling in heart failure-adapt or die. Nat Med. 2008;14:485–7. - PubMed
-
- Post SR, Hammond HK, Insel PA. Beta-adrenergic receptors and receptor signaling in heart failure. Annu Rev Pharmacol Toxicol. 1999;39:343–60. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
