

Molecular Basis for Lytic Bacteriophage Resistance in Enterococci
- PMID: 27578757
- PMCID: PMC4999554
- DOI: 10.1128/mBio.01304-16
Molecular Basis for Lytic Bacteriophage Resistance in Enterococci
Abstract
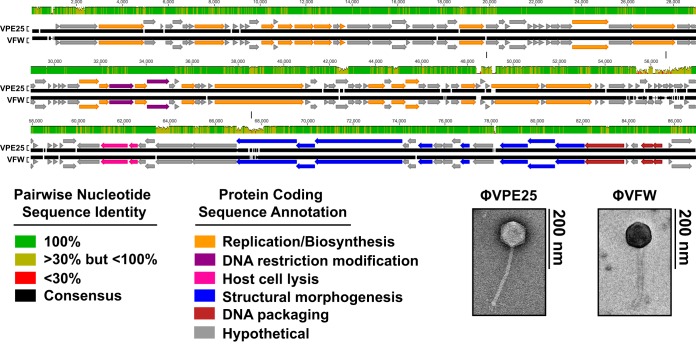
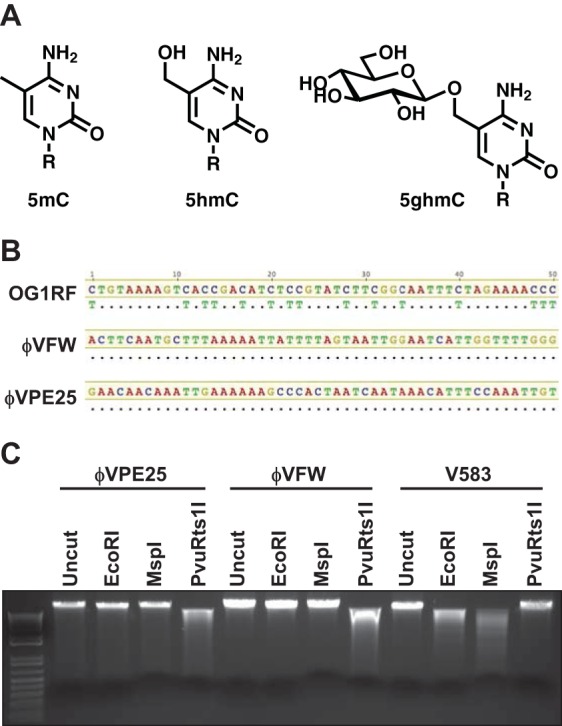
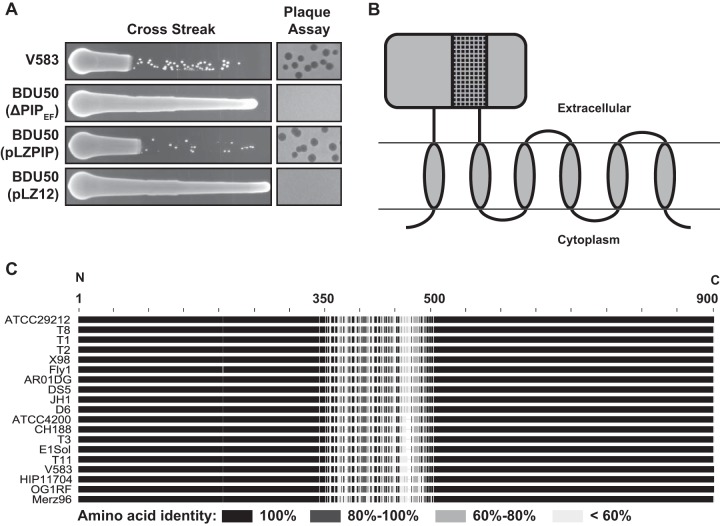
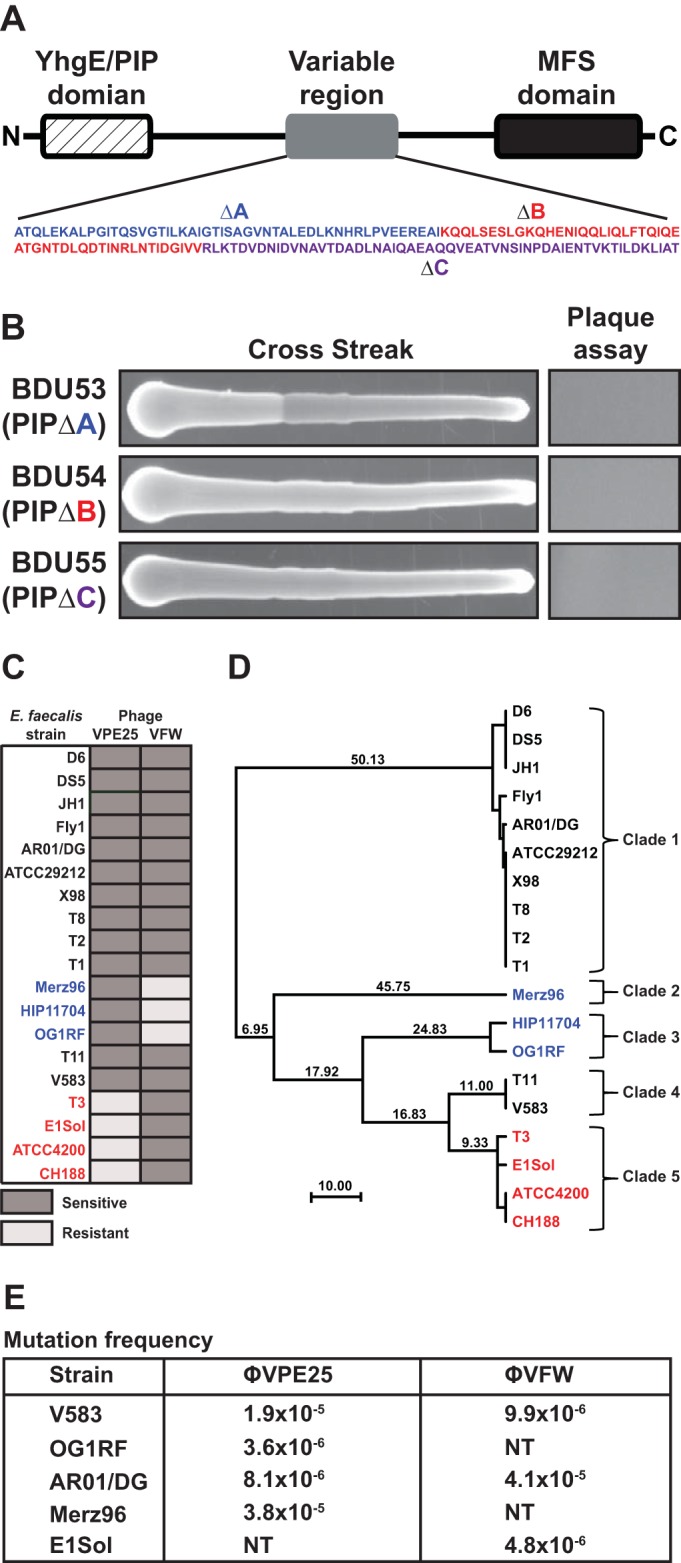
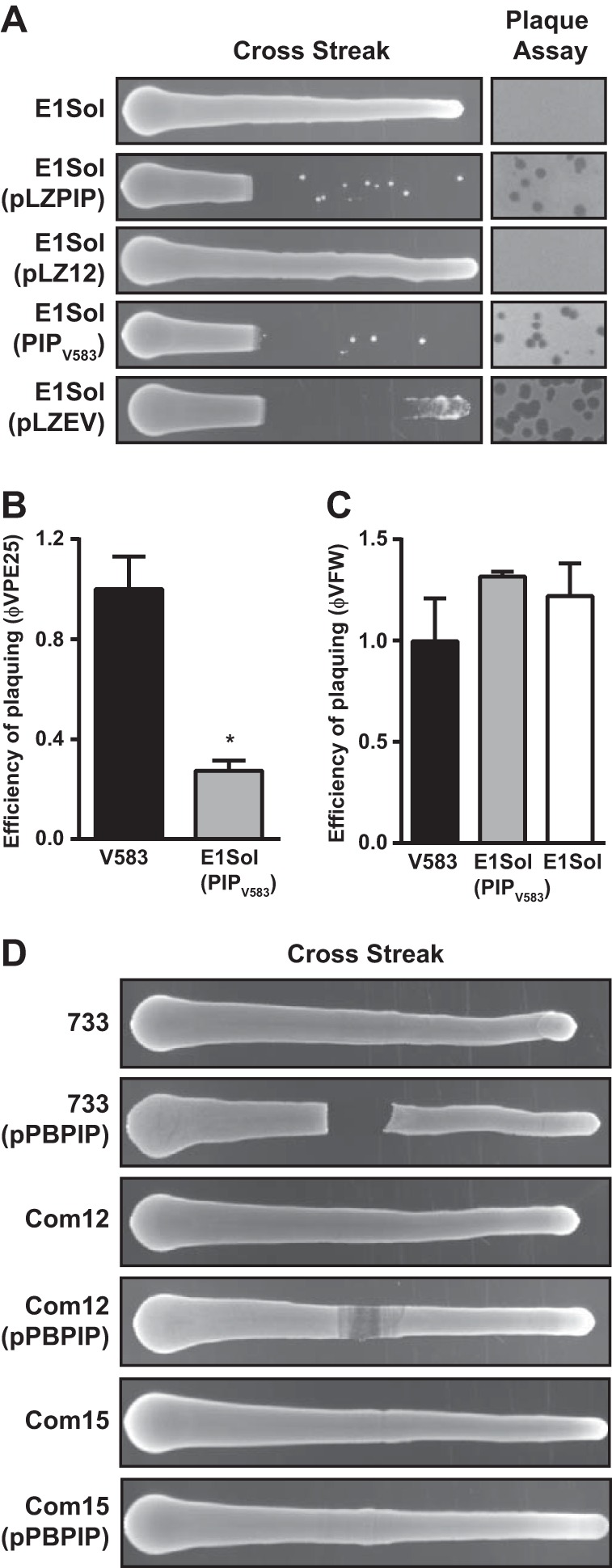
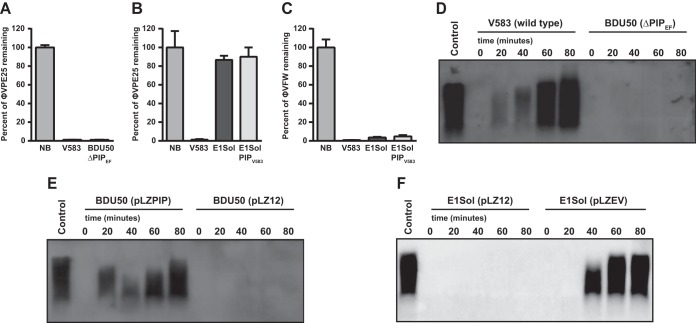
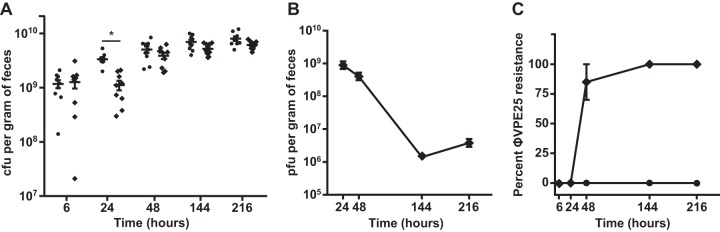
The human intestine harbors diverse communities of bacteria and bacteriophages. Given the specificity of phages for their bacterial hosts, there is growing interest in using phage therapies to combat the rising incidence of multidrug-resistant bacterial infections. A significant barrier to such therapies is the rapid development of phage-resistant bacteria, highlighting the need to understand how bacteria acquire phage resistance in vivo Here we identify novel lytic phages in municipal raw sewage that kill Enterococcus faecalis, a Gram-positive opportunistic pathogen that resides in the human intestine. We show that phage infection of E. faecalis requires a predicted integral membrane protein that we have named PIPEF (for phage infection protein from E. faecalis). We find that PIPEF is conserved in E. faecalis and harbors a 160-amino-acid hypervariable region that determines phage tropism for distinct enterococcal strains. Finally, we use a gnotobiotic mouse model of in vivo phage predation to show that the sewage phages temporarily reduce E. faecalis colonization of the intestine but that E. faecalis acquires phage resistance through mutations in PIPEF Our findings define the molecular basis for an evolutionary arms race between E. faecalis and the lytic phages that prey on them. They also suggest approaches for engineering E. faecalis phages that have altered host specificity and that can subvert phage resistance in the host bacteria.
Importance: Bacteriophage therapy has received renewed attention as a potential solution to the rise in antibiotic-resistant bacterial infections. However, bacteria can acquire phage resistance, posing a major barrier to phage therapy. To overcome this problem, it is necessary to understand phage resistance mechanisms in bacteria. We have unraveled one such resistance mechanism in Enterococcus faecalis, a Gram-positive natural resident of the human intestine that has acquired antibiotic resistance and can cause opportunistic infections. We have identified a cell wall protein hypervariable region that specifies phage tropism in E. faecalis Using a gnotobiotic mouse model of in vivo phage predation, we show that E. faecalis acquires phage resistance through mutations in this cell wall protein. Our findings define the molecular basis for lytic phage resistance in E. faecalis They also suggest opportunities for engineering E. faecalis phages that circumvent the problem of bacterial phage resistance.
Copyright © 2016 Duerkop et al.
Figures
Similar articles
-
Bacteriophage Resistance Alters Antibiotic-Mediated Intestinal Expansion of Enterococci.Infect Immun. 2019 May 21;87(6):e00085-19. doi: 10.1128/IAI.00085-19. Print 2019 Jun. Infect Immun. 2019. PMID: 30936157 Free PMC article.
-
Characterization and Anti-Biofilm Activity of Lytic Enterococcus Phage vB_Efs8_KEN04 against Clinical Isolates of Multidrug-Resistant Enterococcus faecalis in Kenya.Viruses. 2024 Aug 9;16(8):1275. doi: 10.3390/v16081275. Viruses. 2024. PMID: 39205249 Free PMC article.
-
Therapeutic potential of a newly isolated bacteriophage against multi-drug resistant Enterococcus faecalis infections: in vitro and in vivo characterization.BMC Microbiol. 2025 Feb 20;25(1):80. doi: 10.1186/s12866-025-03785-z. BMC Microbiol. 2025. PMID: 39979834 Free PMC article.
-
Evaluation of Phage Therapy in the Context of Enterococcus faecalis and Its Associated Diseases.Viruses. 2019 Apr 20;11(4):366. doi: 10.3390/v11040366. Viruses. 2019. PMID: 31010053 Free PMC article. Review.
-
Molecular mechanisms of enterococcal-bacteriophage interactions and implications for human health.Curr Opin Microbiol. 2020 Aug;56:38-44. doi: 10.1016/j.mib.2020.06.003. Epub 2020 Jul 8. Curr Opin Microbiol. 2020. PMID: 32652484 Free PMC article. Review.
Cited by
-
Enterococcus faecalis Countermeasures Defeat a Virulent Picovirinae Bacteriophage.Viruses. 2019 Jan 10;11(1):48. doi: 10.3390/v11010048. Viruses. 2019. PMID: 30634666 Free PMC article.
-
The Novel Enterococcus Phage vB_EfaS_HEf13 Has Broad Lytic Activity Against Clinical Isolates of Enterococcus faecalis.Front Microbiol. 2019 Dec 17;10:2877. doi: 10.3389/fmicb.2019.02877. eCollection 2019. Front Microbiol. 2019. PMID: 31921055 Free PMC article.
-
Lytic bacteriophages facilitate antibiotic sensitization of Enterococcus faecium.Antimicrob Agents Chemother. 2023 May 1;65(5):e00143-21. doi: 10.1128/AAC.00143-21. Epub 2021 Mar 1. Antimicrob Agents Chemother. 2023. PMID: 33649110 Free PMC article.
-
Vancomycin Resistance in Enterococcus and Staphylococcus aureus.Microorganisms. 2022 Dec 21;11(1):24. doi: 10.3390/microorganisms11010024. Microorganisms. 2022. PMID: 36677316 Free PMC article. Review.
-
Enterococcal biofilm-A nidus for antibiotic resistance transfer?J Appl Microbiol. 2022 May;132(5):3444-3460. doi: 10.1111/jam.15441. Epub 2022 Jan 26. J Appl Microbiol. 2022. PMID: 34990042 Free PMC article. Review.
References
-
- Lebreton F, Willems RJL, Gilmore MS. 2014. Enterococcus diversity, origins in nature, and gut colonization. In Gilmore MS, Clewell DB, Ike Y, Shankar N (ed), Enterococci: from commensals to leading causes of drug resistant infection. Massachusetts Eye and Ear Infirmary, Boston, MA. - PubMed
-
- Ubeda C, Taur Y, Jenq RR, Equinda MJ, Son T, Samstein M, Viale A, Socci ND, van den Brink MR, Kamboj M, Pamer EG. 2010. Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J Clin Invest 120:4332–4341. doi: 10.1172/JCI43918. - DOI - PMC - PubMed
-
- Sievert DM, Ricks P, Edwards JR, Schneider A, Patel J, Srinivasan A, Kallen A, Limbago B, Fridkin S, National Healthcare Safety Network (NHSN) Team and Participating NHSN Facilities . 2013. Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect Control Hosp Epidemiol 34:1–14. doi: 10.1086/668770. - DOI - PubMed
-
- Agudelo Higuita NI, Huycke MM. 2014. Enterococcal disease, epidemiology, and implications for treatment. In Gilmore MS, Clewell DB, Ike Y, Shankar N (ed), Enterococci: from commensals to leading causes of drug resistant infection. Massachusetts Eye and Ear Infirmary, Boston, MA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
