

Interactions of Prototype Foamy Virus Capsids with Host Cell Polo-Like Kinases Are Important for Efficient Viral DNA Integration
- PMID: 27579920
- PMCID: PMC5006980
- DOI: 10.1371/journal.ppat.1005860
Interactions of Prototype Foamy Virus Capsids with Host Cell Polo-Like Kinases Are Important for Efficient Viral DNA Integration
Erratum in
-
Correction: Interactions of Prototype Foamy Virus Capsids with Host Cell Polo-Like Kinases Are Important for Efficient Viral DNA Integration.PLoS Pathog. 2016 Oct 10;12(10):e1005956. doi: 10.1371/journal.ppat.1005956. eCollection 2016 Oct. PLoS Pathog. 2016. PMID: 27723818 Free PMC article.
Abstract
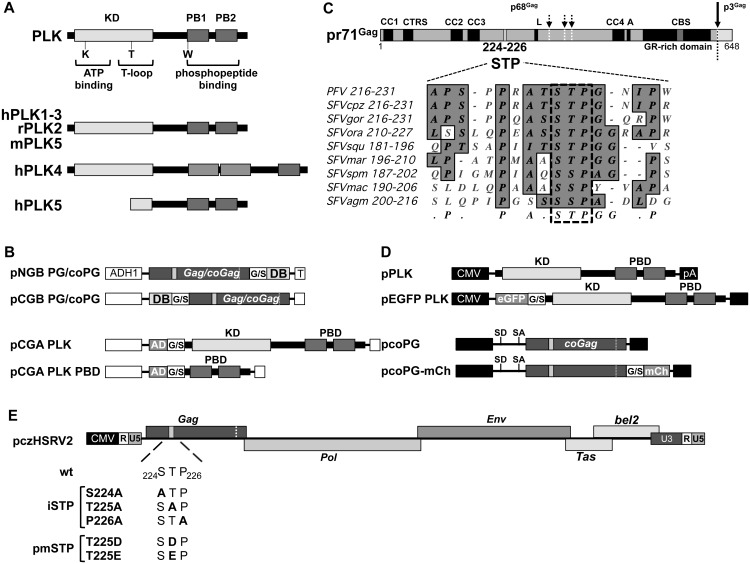
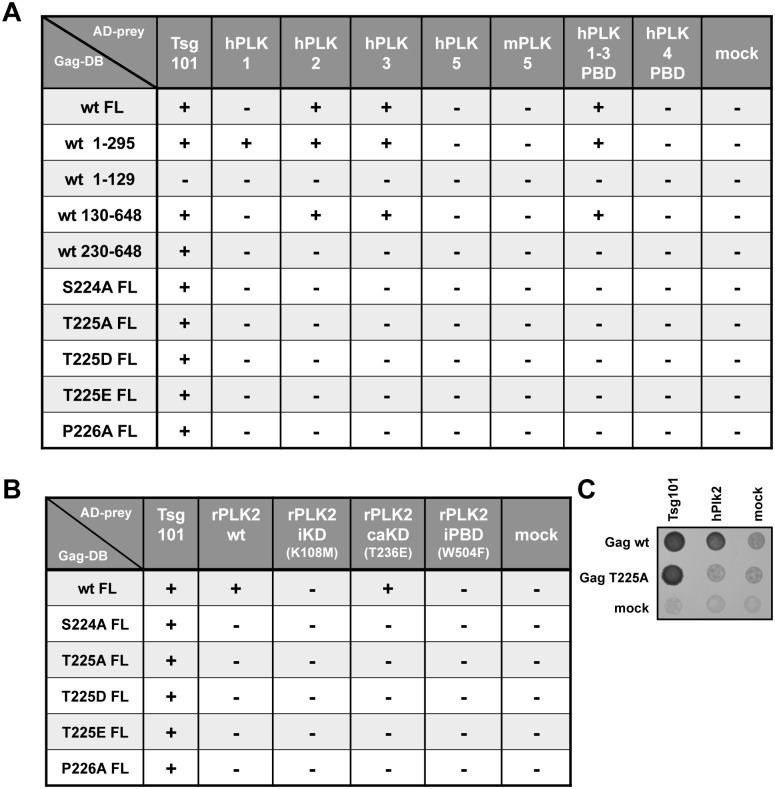
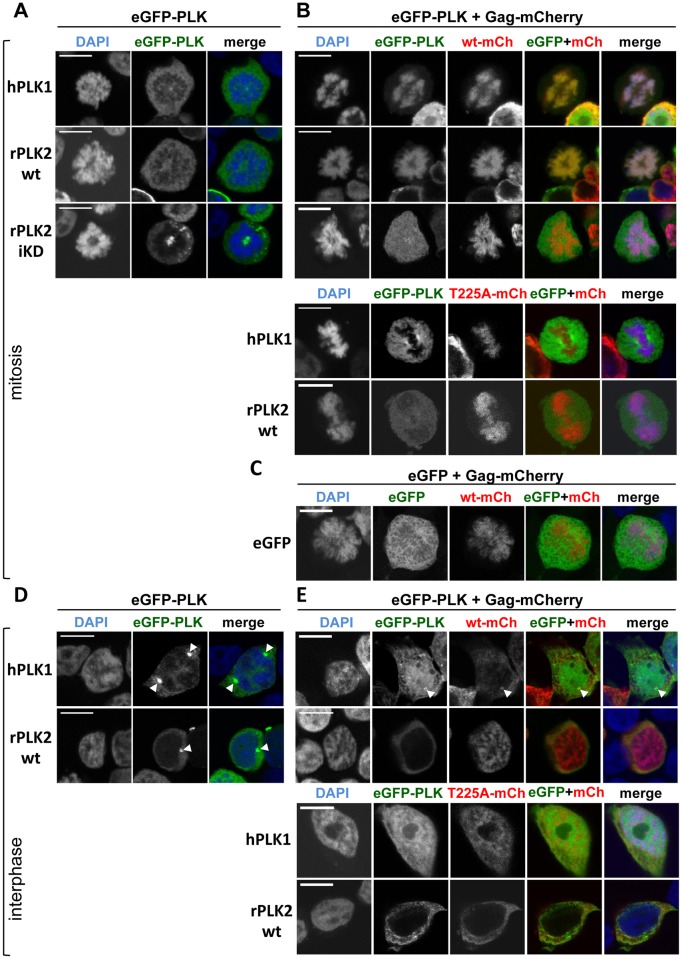
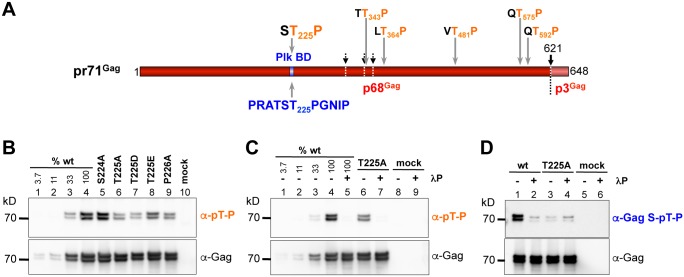
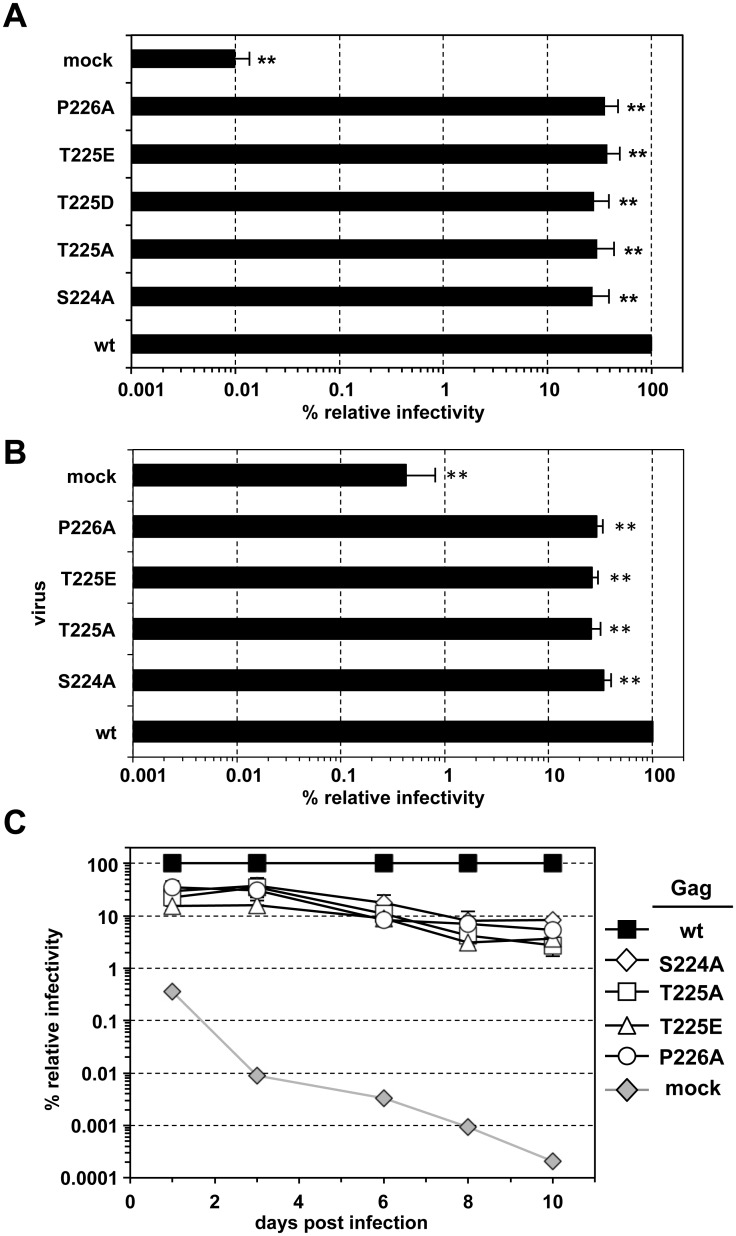
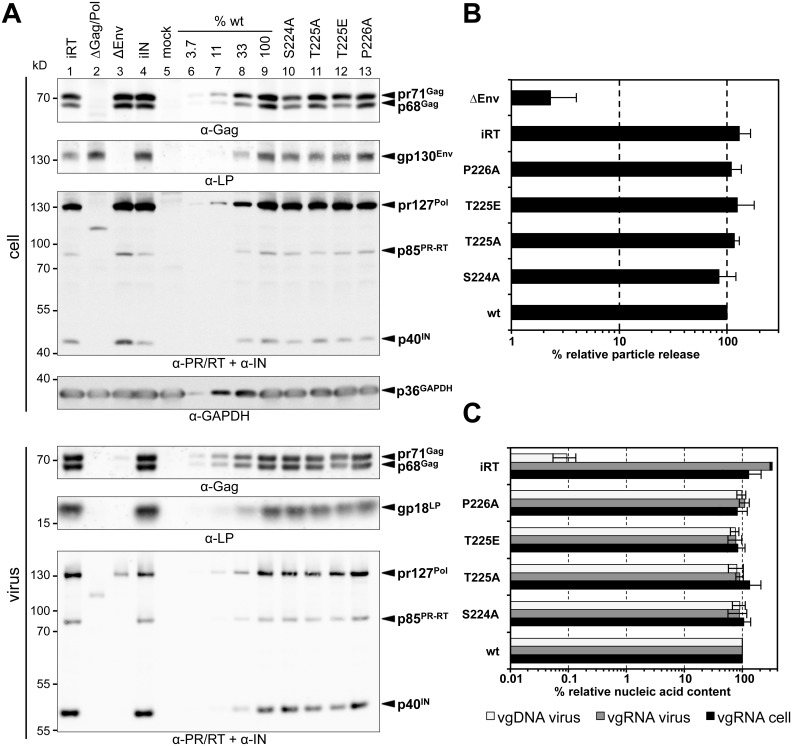
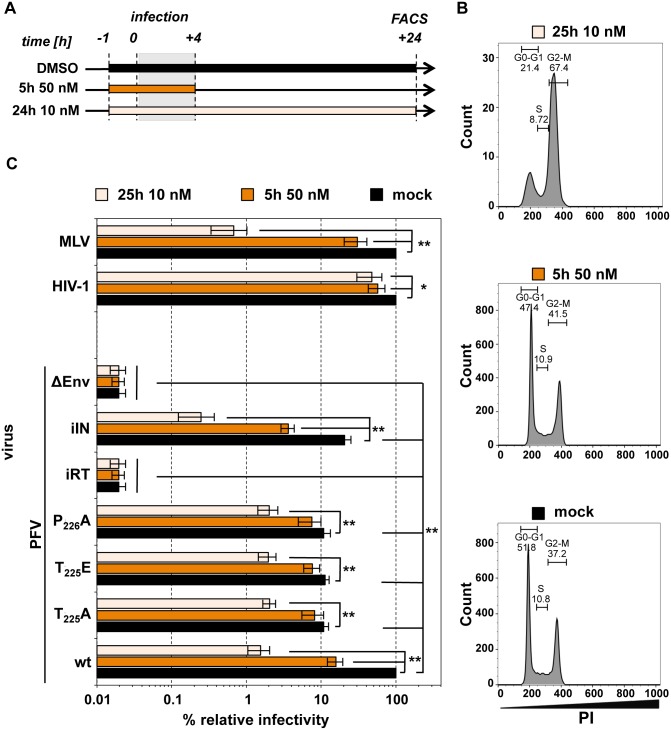
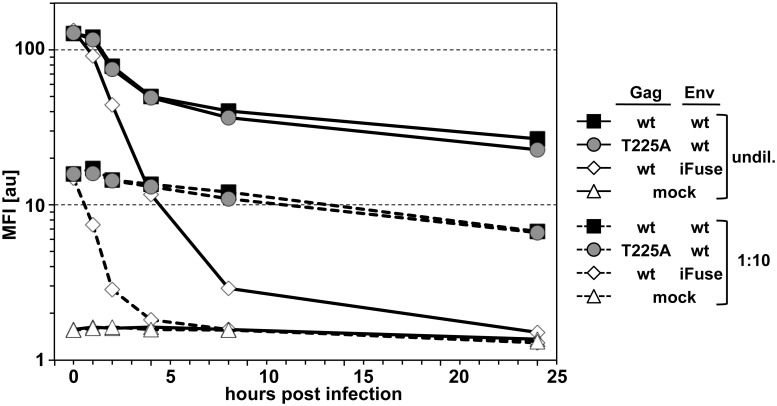
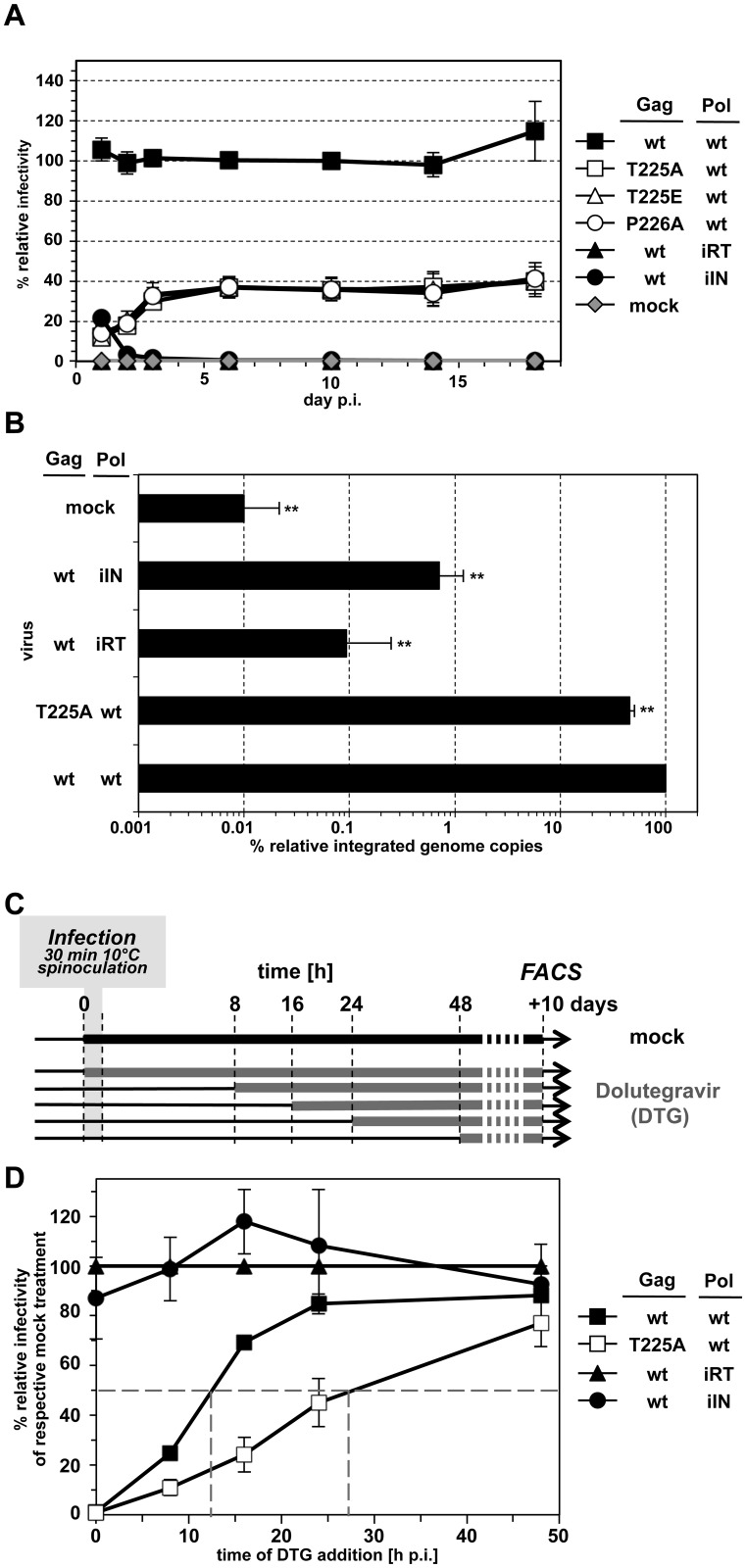
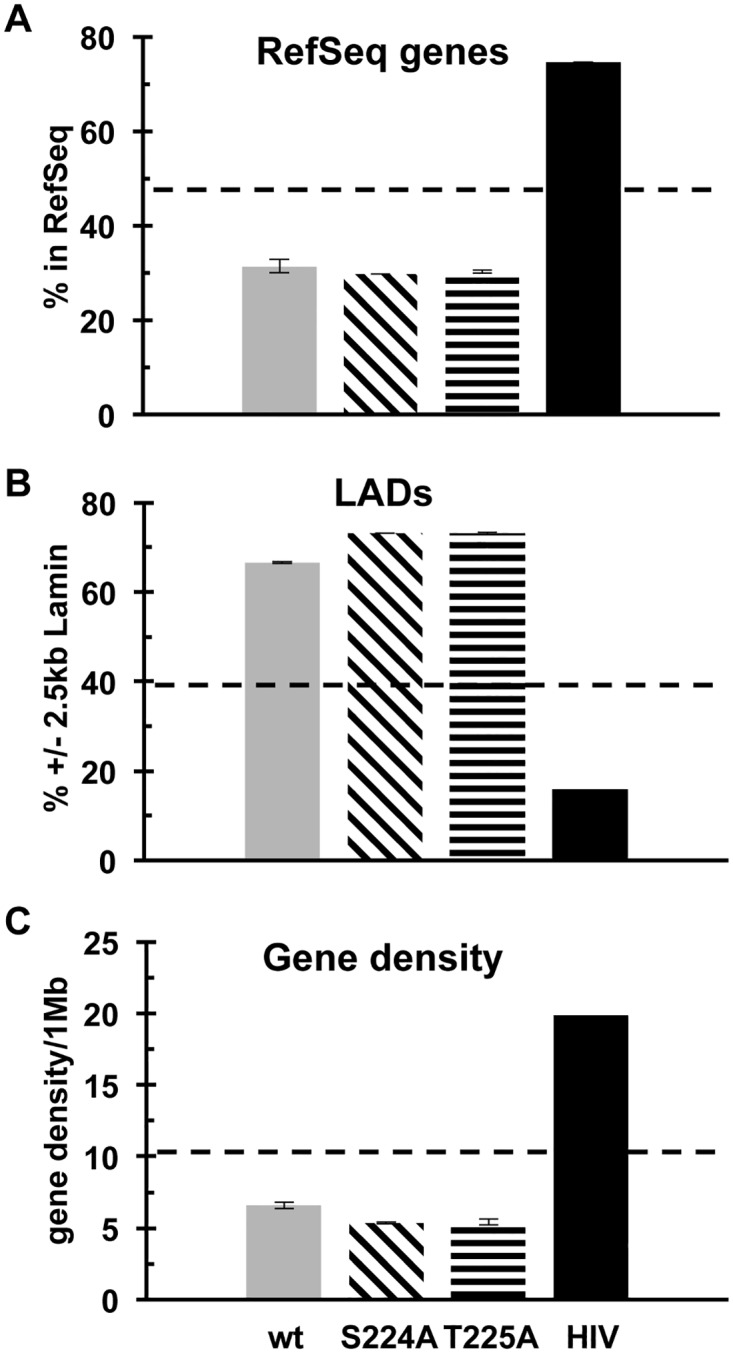
Unlike for other retroviruses, only a few host cell factors that aid the replication of foamy viruses (FVs) via interaction with viral structural components are known. Using a yeast-two-hybrid (Y2H) screen with prototype FV (PFV) Gag protein as bait we identified human polo-like kinase 2 (hPLK2), a member of cell cycle regulatory kinases, as a new interactor of PFV capsids. Further Y2H studies confirmed interaction of PFV Gag with several PLKs of both human and rat origin. A consensus Ser-Thr/Ser-Pro (S-T/S-P) motif in Gag, which is conserved among primate FVs and phosphorylated in PFV virions, was essential for recognition by PLKs. In the case of rat PLK2, functional kinase and polo-box domains were required for interaction with PFV Gag. Fluorescently-tagged PFV Gag, through its chromatin tethering function, selectively relocalized ectopically expressed eGFP-tagged PLK proteins to mitotic chromosomes in a Gag STP motif-dependent manner, confirming a specific and dominant nature of the Gag-PLK interaction in mammalian cells. The functional relevance of the Gag-PLK interaction was examined in the context of replication-competent FVs and single-round PFV vectors. Although STP motif mutated viruses displayed wild type (wt) particle release, RNA packaging and intra-particle reverse transcription, their replication capacity was decreased 3-fold in single-cycle infections, and up to 20-fold in spreading infections over an extended time period. Strikingly similar defects were observed when cells infected with single-round wt Gag PFV vectors were treated with a pan PLK inhibitor. Analysis of entry kinetics of the mutant viruses indicated a post-fusion defect resulting in delayed and reduced integration, which was accompanied with an enhanced preference to integrate into heterochromatin. We conclude that interaction between PFV Gag and cellular PLK proteins is important for early replication steps of PFV within host cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
SGK1, a Serine/Threonine Kinase, Inhibits Prototype Foamy Virus Replication.Microbiol Spectr. 2022 Jun 29;10(3):e0199521. doi: 10.1128/spectrum.01995-21. Epub 2022 Apr 19. Microbiol Spectr. 2022. PMID: 35438526 Free PMC article.
-
Correct capsid assembly mediated by a conserved YXXLGL motif in prototype foamy virus Gag is essential for infectivity and reverse transcription of the viral genome.J Virol. 2007 Apr;81(7):3317-26. doi: 10.1128/JVI.01866-06. Epub 2007 Jan 17. J Virol. 2007. PMID: 17229703 Free PMC article.
-
Orthoretroviral-like prototype foamy virus Gag-Pol expression is compatible with viral replication.Retrovirology. 2011 Aug 15;8:66. doi: 10.1186/1742-4690-8-66. Retrovirology. 2011. PMID: 21843316 Free PMC article.
-
Foamy virus assembly with emphasis on pol encapsidation.Viruses. 2013 Mar 20;5(3):886-900. doi: 10.3390/v5030886. Viruses. 2013. PMID: 23518575 Free PMC article. Review.
-
The Unique, the Known, and the Unknown of Spumaretrovirus Assembly.Viruses. 2021 Jan 13;13(1):105. doi: 10.3390/v13010105. Viruses. 2021. PMID: 33451128 Free PMC article. Review.
Cited by
-
Correction: Interactions of Prototype Foamy Virus Capsids with Host Cell Polo-Like Kinases Are Important for Efficient Viral DNA Integration.PLoS Pathog. 2016 Oct 10;12(10):e1005956. doi: 10.1371/journal.ppat.1005956. eCollection 2016 Oct. PLoS Pathog. 2016. PMID: 27723818 Free PMC article.
-
TraFo-CRISPR: Enhanced Genome Engineering by Transient Foamy Virus Vector-Mediated Delivery of CRISPR/Cas9 Components.Mol Ther Nucleic Acids. 2019 Dec 6;18:708-726. doi: 10.1016/j.omtn.2019.10.006. Epub 2019 Oct 17. Mol Ther Nucleic Acids. 2019. PMID: 31726388 Free PMC article.
-
Master mitotic kinases regulate viral genome delivery during papillomavirus cell entry.Nat Commun. 2023 Jan 23;14(1):355. doi: 10.1038/s41467-023-35874-w. Nat Commun. 2023. PMID: 36683055 Free PMC article.
-
The invariant arginine within the chromatin-binding motif regulates both nucleolar localization and chromatin binding of Foamy virus Gag.Retrovirology. 2018 Jul 11;15(1):48. doi: 10.1186/s12977-018-0428-z. Retrovirology. 2018. PMID: 29996845 Free PMC article.
-
Polo-Like Kinase 2: From Principle to Practice.Front Oncol. 2022 Jul 8;12:956225. doi: 10.3389/fonc.2022.956225. eCollection 2022. Front Oncol. 2022. PMID: 35898867 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
