

Multiciliated Cells in Animals
- PMID: 27580630
- PMCID: PMC5131768
- DOI: 10.1101/cshperspect.a028233
Multiciliated Cells in Animals
Abstract
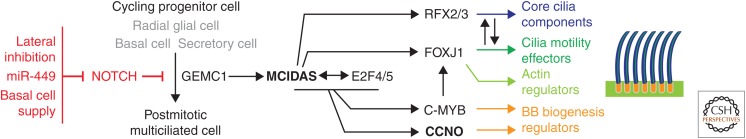
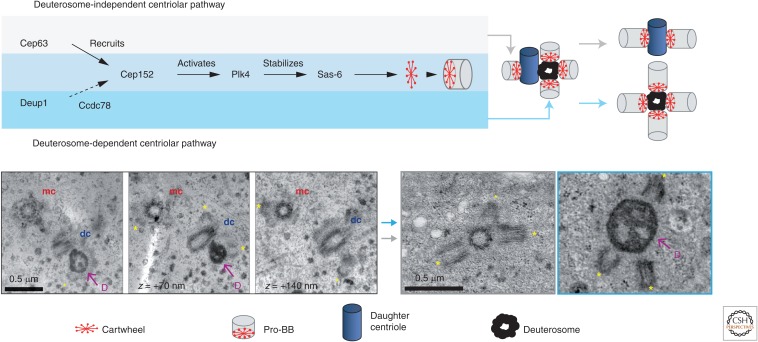
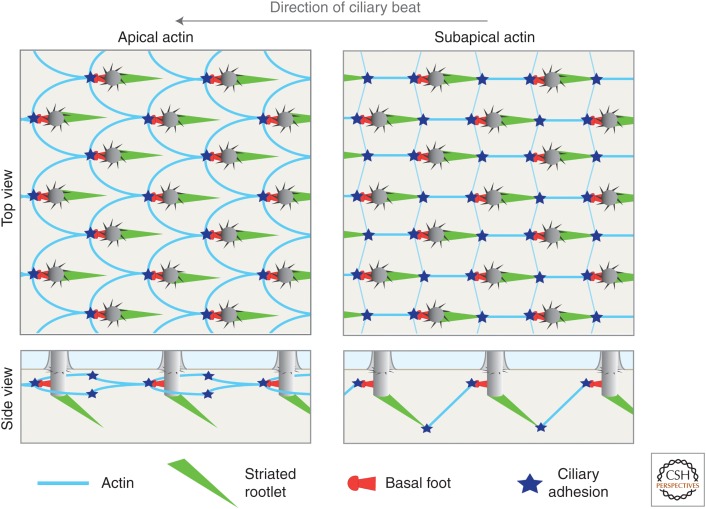
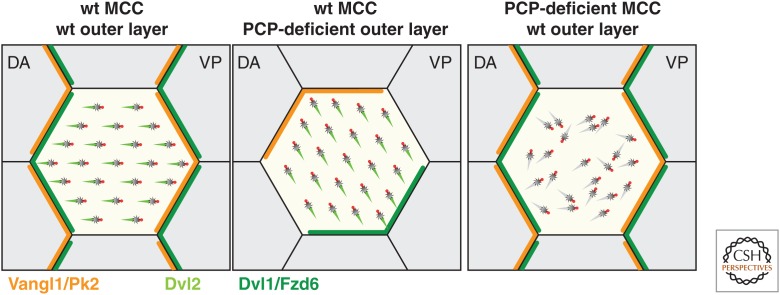
Many animal cells assemble single cilia involved in motile and/or sensory functions. In contrast, multiciliated cells (MCCs) assemble up to 300 motile cilia that beat in a coordinate fashion to generate a directional fluid flow. In the human airways, the brain, and the oviduct, MCCs allow mucus clearance, cerebrospinal fluid circulation, and egg transportation, respectively. Impairment of MCC function leads to chronic respiratory infections and increased risks of hydrocephalus and female infertility. MCC differentiation during development or repair involves the activation of a regulatory cascade triggered by the inhibition of Notch activity in MCC progenitors. The downstream events include the simultaneous assembly of a large number of basal bodies (BBs)-from which cilia are nucleated-in the cytoplasm of the differentiating MCCs, their migration and docking at the plasma membrane associated to an important remodeling of the actin cytoskeleton, and the assembly and polarization of motile cilia. The direction of ciliary beating is coordinated both within cells and at the tissue level by a combination of planar polarity cues affecting BB position and hydrodynamic forces that are both generated and sensed by the cilia. Herein, we review the mechanisms controlling the specification and differentiation of MCCs and BB assembly and organization at the apical surface, as well as ciliary assembly and coordination in MCCs.
Copyright © 2016 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Al Jord A, Lemaitre AI, Delgehyr N, Faucourt, Spassky N, Meunier A. 2014. Centriole amplification by mother and daughter centrioles differs in multiciliated cells. Nature 516: 104–107. - PubMed
-
- Amirav I, Wallmeier J, Loges NT, Menchen T, Pennekamp P, Mussaffi H, Abitbul R, Avital A, Bentur L, Dougherty GW, et al. 2016. Systematic analysis of CCNO variants in a defined population: Implications for clinical phenotype and differential diagnosis. Hum Mutat 37: 396–405. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources